The Cyklický AMP nebo adenosin 3 ', 5'-monofosfát je cyklický nukleotid, který funguje jako sekundární posel a je součástí základních prvků biochemické kontroly a intracelulární komunikace v mnoha živých organismech.
Jeho existenci prokázali před téměř 70 lety Sutherland a Rall (1958), kteří popsali fenomén akumulace tohoto nukleotidu v jaterních buňkách v důsledku podávání epinefrinu (adrenalinu).
Od svého objevu je cyklický AMP spojován u savců s působením mnoha hormonů, s endokrinní a exokrinní sekrecí, s uvolňováním neurotransmiterů v synapse a neuromuskulárních spojích, mezi mnoha dalšími funkcemi..
Jeho syntéza je katalyzována třemi rodinami proteinů asociovaných s plazmatickou membránou známých jako adenylcykláza nebo adenylátcykláza, které jsou schopné produkovat cyklickou sloučeninu z ATP a uvolňovat pyrofosfát do buňky..
Jeho degradace je naopak zprostředkována enzymy z rodiny fosfodiesterázy, což jsou rozpustné proteiny vyskytující se hlavně v cytosolu.
Tyto enzymy, a proto cyklický AMP, se nacházejí v extrémně rozmanitých organismech, tak jednoduchých jako jednobuněčné řasy a mnoho dalších mikroorganismů (bakterie a další) a tak složitých jako mnohobuněčná zvířata se složitými signálními cestami..
Ačkoli je jeho přítomnost v rostlinách předmětem diskuse, existují určité důkazy, které naznačují, že některé druhy rostlin mají aktivitu adenylátcyklázy, i když její funkce nebyla uspokojivě stanovena..
Rejstřík článků
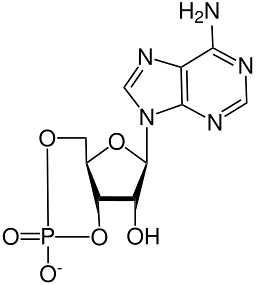
Chemická struktura cyklického AMP byla objasněna rentgenovou krystalografií a studiemi protonové nukleární magnetické rezonance..
Toto je tepelně stabilní cyklická molekula ("tepelně stabilní “) a stabilnější vůči alkalické hydrolýze než jeho necyklický protějšek, AMP nebo adenosinmonofosfát.
Stejně jako všechny fosfátové nukleotidy má cyklický AMP fosfátovou skupinu připojenou k uhlíkovému kyslíku v poloze 5 'molekuly ribózy, která je zase připojena k dusíkaté bázi heterocyklického kruhu prostřednictvím uhlíku v poloze 1', což odpovídá adenin.
Fosfátová skupina ribózového cukru je na rozdíl od necyklických fosfátových nukleotidů fúzována na trans fosfodiesterovou vazbou s kyslíky uhlíků v poloze 3 'a 5' ribózy (3 ', 5'-trans-tavený fosfát).
Toto spojení omezuje pohyb furanového prstence, který tvoří ribózu, a uzavírá fosfátovou skupinu v „křeslové“ konformaci..
Ve srovnání s necyklickými nukleotidy jsou cyklické AMP a další příbuzné nukleotidy menší molekuly s nižší polaritou, což je důležitý faktor pro jejich diferenciaci proteiny, které na ně reagují..
Konformace glykosidové vazby mezi ribózou a adeninovým kruhem má určitou volnost otáčení. To je také důležitý strukturální parametr pro jeho odlišení od jiných nukleotidů (nejen identita dusíkaté báze)..
Jako sekundární posel se cyklický AMP účastní aktivace mnoha signálních procesů (po jejich syntéze) nebo aktivace různých enzymů „po proudu“ v signální kaskádě, kterou je produkován..
Podílí se na jaterní glykogenolýze a na uvolňování inzulínu z pankreatu, na uvolňování amylázy ze slinných žláz a na působení estrogenu v děloze.
Má univerzální funkce při kontrole genové exprese a při integraci více metabolických funkcí. Mnoho cytokinů používá k výkonu svých funkcí jak vápník, tak cyklický AMP..
Mezi hormony, které používají cyklický AMP ve svých signálních procesech (buď zvýšením nebo snížením jeho intracelulární koncentrace), lze mimo jiné zahrnout katecholaminy, glukagon, vasopresin, paratyroidní hormon, prostaglandiny, inzulín, melatonin a adrenalin.
Další z jeho mnoha funkcí je inhibovat růst, diferenciaci a proliferaci T buněk u savců, pravděpodobně aktivací nebo indukcí represoru regulačních cytokinů těchto procesů v takových buňkách..
Cyklický AMP a adenylátcyklázy, které jej produkují, také souvisí s funkcí mnoha receptorů proteinu spojených s G proteinem, které jsou spojeny s různými signálními mechanismy a dalšími důležitými buněčnými procesy..
U savců má cyklický AMP funkce v regulaci glykolytické a glukoneogenní dráhy prostřednictvím inhibice aktivity enzymu fosfofruktokinázy 2 (PFK-2), který katalyzuje druhou reakci glykolýzy..
Tento mechanismus zahrnuje účast hormonu glukagonu na aktivaci jaterní adenylátcyklázy, což způsobuje značné zvýšení koncentrace cyklického AMP..
Tento cyklický AMP aktivuje proteinkinázu závislou na cAMP, která fosforyluje a inhibuje aktivitu fosfofruktokinázy PFK-2, což je bifunkční enzym s aktivitou fruktózo-bisfosfatázy..
První posel (proměnlivé chemické povahy), který dosáhne specifické buňky jako externí stimul, interaguje s enzymem adenylátcyklázy v plazmatické membráně a indukuje produkci cyklického AMP.
Zvýšení koncentrace cyklických funkcí AMP při aktivaci dalších faktorů (obecně enzymatických), které mají mimo jiné další funkce při inhibici nebo aktivaci metabolických procesů nebo genové transkripce..
Jednou z hlavních funkcí spojených s touto regulační molekulou je aktivace fosforylázových nebo kinázových enzymů, které katalyzují přidání nebo odstranění fosforylových skupin k jiným proteinům a enzymům..
Za normálních okolností je excitace buňky doprovázena zvýšením koncentrace cyklického AMP, souběžně se zvýšením transportu vápníku směrem k buňce, která má funkce při aktivaci cyklických enzymů adenylcyklázy produkujících AMP..
Syntéza i přenos zprávy a degradace cyklického AMP v buňkách jsou jemně regulované procesy, které se podílejí na udržování tělesné homeostázy..


Zatím žádné komentáře