The glykolipidy Jsou to membránové lipidy se sacharidy v jejich skupinách polárních hlav. Představují nejvíce asymetrickou distribuci mezi membránovými lipidy, protože se nacházejí výlučně ve vnější monovrstvě buněčných membrán, přičemž jsou obzvláště hojné v plazmatické membráně.
Stejně jako většina membránových lipidů mají glykolipidy hydrofobní oblast složenou z apolárních uhlovodíkových ocasů a hlavu nebo polární oblast, která může být tvořena různými třídami molekul, v závislosti na daném glykolipidu..
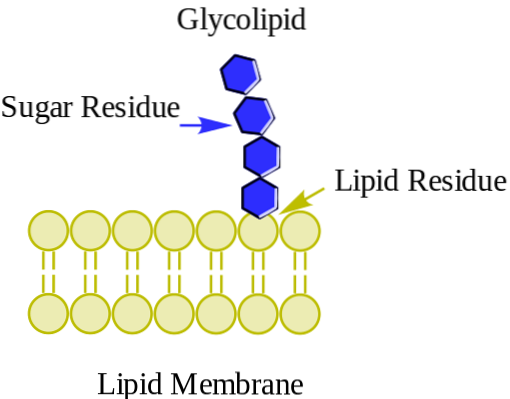
Glykolipidy lze nalézt v jednobuněčných organismech, jako jsou bakterie a kvasinky, stejně jako v komplexních organismech jako zvířata a rostliny.
V živočišných buňkách jsou glykolipidy převážně složeny ze sfingosinového skeletu, zatímco v rostlinách dva nejčastější odpovídají diglyceridům a derivátům kyseliny sulfonové. V bakteriích jsou také glykosylglyceridy a deriváty acylovaných cukrů.
V rostlinách jsou glykolipidy koncentrovány v chloroplastických membránách, zatímco u zvířat jsou hojné v plazmatické membráně. Spolu s glykoproteiny a proteoglykany jsou glykolipidy důležitou součástí glykokalyxu, což je zásadní pro mnoho buněčných procesů.
Glykolipidy, zejména živočišné buňky, mají tendenci se navzájem sdružovat prostřednictvím vodíkové vazby mezi svými sacharidovými skupinami a van der Waalsovými silami mezi jejich řetězci mastných kyselin. Tyto lipidy jsou přítomny v membránových strukturách známých jako lipidové rafty, které mají více funkcí.
Funkce glykolipidů jsou různé, ale u eukaryot je jejich umístění na vnější straně plazmatické membrány relevantní z různých hledisek, zejména v procesech komunikace, adheze a diferenciace buněk..
Rejstřík článků
Glykolipidy jsou glykokonjugáty, které tvoří velmi heterogenní skupinu molekul, jejichž společnou charakteristikou je přítomnost sacharidových zbytků vázaných glukosidickými vazbami na hydrofobní část, kterou může být acyl-glycerol, ceramid nebo prenyl fosfát.
Jeho klasifikace je založena na molekulárním skeletu, který je spojnicí mezi hydrofobní a polární oblastí. V závislosti na identitě této skupiny tedy máme:
Tyto glykolipidy, jako glycerolipidy, mají diacylglycerol nebo monoalkyl-monoacylglycerol, hlavní řetězec, ke kterému jsou zbytky cukru připojeny glukosidickými vazbami..
Glykoglycerolipidy jsou relativně jednotné, pokud jde o jejich složení sacharidů, a zbytky galaktózy nebo glukózy lze nalézt v jejich struktuře, od které je odvozena jejich hlavní klasifikace, a to:
Tyto lipidy mají jako „kostru“ molekulu část ceramidu, která může mít připojené různé molekuly mastných kyselin..
Jsou to vysoce variabilní lipidy, a to nejen z hlediska složení jejich hydrofobních řetězců, ale také z hlediska zbytků sacharidů v jejich polární hlavě. Jsou hojné v mnoha savčích tkáních.
Jejich klasifikace je založena spíše na typu substituce nebo identitě sacharidové části než na oblasti složené z hydrofobních řetězců. Podle typů substituce je klasifikace těchto sfingolipidů následující:
Neutrální glukosfingolipidy: ty, které obsahují v sacharidové části hexózy, N-acetylhexosaminy a methylpentózy.
Sulfatidy: jsou to glukosfingolipidy, které obsahují sulfátové estery. Jsou záporně nabité a jsou obzvláště hojné v myelinových pochvách mozkových buněk. Nejběžnější jsou zbytky galaktózy.
Gangliosidy: také známé jako sialosyl glykolipidy, jsou to ty, které obsahují kyselinu sialovou, a proto jsou také známé jako kyselé glykosfingolipidy.
Fosfoinositido-glykolipidy: kostra je tvořena fosfoinositido-ceramidy.
Jsou to lipidy, které jsou obvykle považovány za stabilní kotvy proteinů v lipidové dvojvrstvě. Jsou posttranslačně přidávány na C-terminální konec mnoha proteinů, které se obvykle nacházejí na vnější straně cytoplazmatické membrány..
Skládají se z centra glukanu, fosfolipidového ocasu a části fosfoethanolaminu, která je spojuje..
Glykolipidy mohou mít sacharidové skupiny připojené k molekule N- nebo O-glukosidovými vazbami, a to i prostřednictvím ne-glukosidových vazeb, jako jsou esterové nebo amidové vazby..
Sacharidová část je vysoce variabilní, nejen ve struktuře, ale také ve složení. Tato sacharidová část může být složena z mono-, di-, oligo- nebo polysacharidů různých typů. Mohou mít amino cukry a dokonce kyselé, jednoduché nebo rozvětvené cukry.
Zde je stručný popis obecné struktury tří hlavních tříd glykolipidů:
Jak již bylo zmíněno dříve, glykoglycerolipidy u zvířat mohou mít galaktózové nebo glukózové zbytky, fosfátované či nikoli. Řetězce mastných kyselin v těchto lipidech obsahují 16 až 20 atomů uhlíku.
V galakto-glycerolipidech dochází k spojení mezi cukrem a lipidovou kostrou prostřednictvím β-glukosidových vazeb mezi C-1 galaktózy a C-3 glycerolu. Další dva uhlíky glycerolu jsou buď esterifikovány mastnými kyselinami, nebo C1 je substituován alkylovou skupinou a C2 acylovou skupinou..
Obvykle je pozorován jediný zbytek galaktózy, i když byla hlášena existence digalaktoglycerolipidů. Když se jedná o slufogalaktoglycerolipid, obvykle se sulfátová skupina nachází na C-3 zbytku galaktózy.
Struktura glycerolipidů je trochu odlišná, zejména pokud jde o počet zbytků glukózy, což může být až 8 zbytků spojených dohromady vazbami typu α (1-6). Molekula glukózy, která přemosťuje lipidovou kostru, je k ní připojena a (1-3) vazbou..
V sulfoglykoglycerolipidech je sulfátová skupina připojena k uhlíku v poloze 6 terminálního zbytku glukózy.
Stejně jako ostatní sfingolipidy jsou glykosfingolipidy odvozeny od L-serinu kondenzovaného s mastnou kyselinou s dlouhým řetězcem, která tvoří sfingoidovou bázi známou jako sfingosin. Když se jiná mastná kyselina váže na uhlík 2 sfingosinu, produkuje se ceramid, který je společnou bází pro všechny sfingolipidy.
V závislosti na typu sfingolipidu jsou tyto složeny ze zbytků D-glukózy, D-galaktózy, N-acetyl-D-galaktosaminu a N-acetylglukosaminu, jakož i kyseliny sialové. Gangliosidy jsou možná nejrozmanitější a nejsložitější, pokud jde o důsledky oligosacharidových řetězců.
V těchto glykolipidech mohou být zbytky glukanového centra (glukosamin a manóza) modifikovány různými způsoby přidáním fosfoethanolaminových skupin a dalších cukrů. Tato odrůda jim poskytuje velkou strukturální složitost, která je důležitá pro jejich zavedení do membrány..
Chloroplasty mnoha řas a vyšších rostlin jsou obohaceny o neutrální galakto-glycerolipidy, které mají podobné vlastnosti jako cerebrosidy u zvířat. Mono- a digalaktolipidy jsou β-vázány na diglyceridovou skupinu, zatímco sulfolipidy jsou odvozeny pouze od α-glukózy..
V bakteriích jsou glykosylglyceridy strukturně analogické se zvířecími fosfoglyceridy, ale obsahují sacharidové zbytky spojené glykosylací v poloze 3 sn-1,2-diglyceridu. Acylované deriváty cukru neobsahují glycerol, ale mastné kyseliny přímo vázané na cukry.
Nejběžnějšími sacharidovými zbytky mezi bakteriálními glykolipidy jsou galaktóza, glukóza a manóza..
U zvířat hrají glykolipidy důležitou roli mimo jiné v buněčné komunikaci, diferenciaci a proliferaci, onkogenezi, elektrickém odpuzování (v případě polárních glykolipidů)..
Jeho přítomnost v mnoha buněčných membránách zvířat, rostlin a mikroorganismů odpovídá za jeho důležitou funkci, která souvisí zejména s vlastnostmi multifunkčních lipidových vorů..
Sacharidová část glykosfingolipidů je určující pro antigenicitu a imunogenicitu buněk, které ji nesou. Může se účastnit mezibuněčných procesů rozpoznávání i buněčných „sociálních“ aktivit..
Galaktoglycerolipidy v rostlinách, vzhledem k jejich relativnímu množství v rostlinných membránách, hrají důležitou roli při vytváření charakteristik membrány, jako je stabilita a funkční aktivita mnoha membránových proteinů..
Role glykolipidů v bakteriích je také různorodá. Některé glykoglycerolipidy jsou potřebné ke zlepšení stability dvojvrstvy. Slouží také jako prekurzory dalších složek membrány a také podporují růst při nedostatku anoxie nebo fosfátů..
Kotvy GPI nebo glukosidylfosfatidylinositoly jsou také přítomny v lipidových raftech, účastní se signální transdukce, patogeneze mnoha parazitických mikroorganismů a orientace apikální membrány.
Lze tedy říci, že obecné funkce glykolipidů, jak u rostlin, zvířat, tak u bakterií, odpovídají stanovení stability a tekutosti membrány; účast na specifických interakcích lipid-protein a rozpoznávání buněk.
1. Abdel-mawgoud, A. M. a Stephanopoulos, G. (2017). Jednoduché glykolipidy mikrobů: Chemie, biologická aktivita a metabolické inženýrství. Syntetická a systémová biotechnologie, 1-17.
2. Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molekulární biologie buňky (6. vydání). New York: Garland Science.
3. Ando, T., Imamura, A., Ishida, H., & Kiso, M. (2007). Syntéza glykolipidů. Výzkum sacharidů, 797-813.
4. Benson, A. (1964). Lipidy rostlinných membrán. Annu. Rev. Plant. Physiol., 15, 1-16.
5. Bronislaw, L., Liau, Y. U. N. H., & Slomiany, A. (1987). Živočišné glykoglycerolipidy. Prog. Lipid Res., 26, 29-51.
6. Holzl, G. a Dormann, P. (2007). Struktura a funkce glykoglycerolipidů v rostlinách a bakteriích. Prog. Lipid Res., 46, 225-243.
7. Honke, K. (2013). Biosyntéza a biologická funkce sulfoglykolipidů. Proc. Jpn. Acad. Ser. B, 89 (4), 129-138.
8. Kanfer, J., & Hakomori, S. (1983). Sfingolipidová biochemie. (D. Hanahan, ed.), Handbook of Lipid Research 3 (1. vyd.).
9. Koynova, R., & Caffrey, M. (1994). Fáze a fázové přechody glykoglycerolipidů. Chemistry and Physics of Lipids, 69, 181-207.
10. Law, J. (1960). Glykolipidy. Výroční recenze, 29, 131–150.
11. Paulick, M. G. a Bertozzi, C. R. (2008). Glykosylfosfatidylinositolová kotva: komplexní membránová kotva. Biochemistry, 47, 6991-7000.



Zatím žádné komentáře