
The plazmatická membrána, Buněčná membrána, plasmalemma nebo cytoplazmatická membrána, je lipidová struktura, která obklopuje a ohraničuje buňky a je nepostradatelnou součástí jejich architektury. Biomembrány mají tu vlastnost, že obklopují určitou strukturu svým zevnějškem. Jeho hlavní funkcí je sloužit jako bariéra.
Kromě toho řídí průchod částic, které mohou vstupovat a vystupovat. Membránové proteiny fungují jako „molekulární brány“ u poměrně náročných bran. Složení membrány má také roli v rozpoznávání buněk.
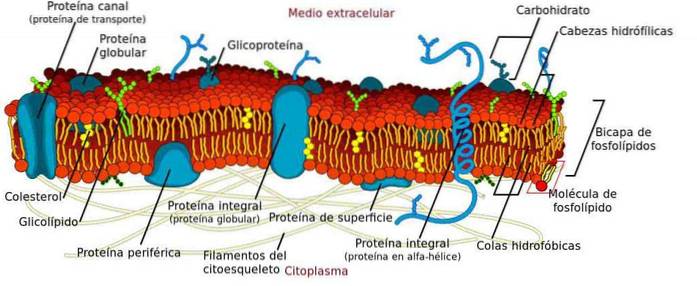
Strukturálně jsou to dvojvrstvy složené z přirozeně uspořádaných fosfolipidů, bílkovin a sacharidů. Podobně fosfolipid představuje fosfor s hlavou a ocasem. Ocas je tvořen uhlíkovými řetězci nerozpustnými ve vodě, které jsou seskupeny dovnitř.
Místo toho jsou hlavy polární a dávají se do vodného buněčného prostředí. Membrány jsou extrémně stabilní konstrukce. Síly, které je udržují, jsou síly van der Waalsa, mezi fosfolipidy, které je tvoří; to jim umožňuje pevně obklopit okraj buněk.
Jsou však také velmi dynamické a plynulé. Vlastnosti membrán se liší podle analyzovaného typu buněk. Například červené krvinky musí být elastické, aby se mohly pohybovat cévami.
Naproti tomu v neuronech má membrána (myelinová pochva) strukturu nezbytnou pro účinné umožnění vedení nervového impulsu..
Rejstřík článků
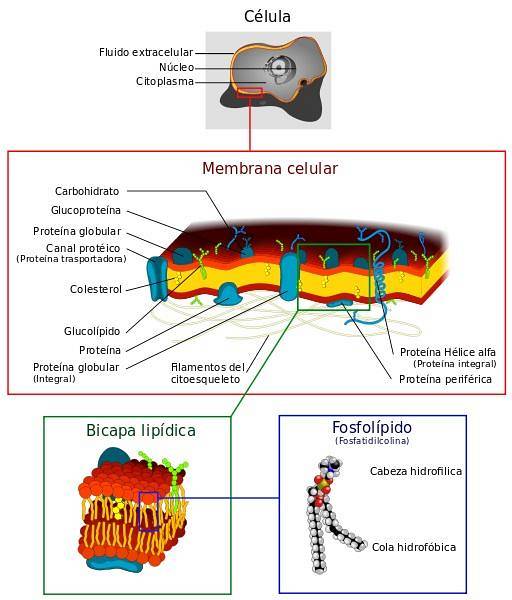
Membrány jsou velmi dynamické struktury, které se velmi liší v závislosti na typu buňky a složení jejích lipidů. Membrány jsou upraveny podle těchto charakteristik následovně:
Membrána není statická entita, chová se jako tekutina. Stupeň tekutosti struktury závisí na několika faktorech, včetně lipidového složení a teploty, které jsou membrány vystaveny..
Když jsou všechny vazby, které existují v uhlíkových řetězcích, nasycené, membrána má tendenci se chovat jako gel a van der Waalsovy interakce jsou stabilní. Naopak, pokud existují dvojné vazby, interakce jsou menší a tekutost se zvyšuje.
Kromě toho existuje účinek délky uhlíkového řetězce. Čím déle to je, tím více interakcí dochází s jeho sousedy, čímž se zvyšuje plynulost. Jak teplota stoupá, zvyšuje se také tekutost membrány.
Cholesterol hraje nepostradatelnou roli při regulaci tekutosti a závisí na koncentraci cholesterolu. Jsou-li fronty dlouhé, působí pro ně cholesterol jako imobilizér, který snižuje tekutost. K tomuto jevu dochází při normální hladině cholesterolu..
Účinek se mění, když jsou hladiny cholesterolu nižší. Interakcí s lipidovými konci je účinek, který způsobuje, jejich oddělení a snížení tekutosti.
Stejně jako tekutost je zakřivení membrány určováno lipidy, které tvoří každou konkrétní membránu..
Zakřivení závisí na velikosti lipidové hlavy a ocasu. Ti, kteří mají dlouhé ocasy a velké hlavy, jsou ploché; ti s relativně menšími hlavami mají sklon k zakřivení mnohem více než předchozí skupina.
Tato vlastnost je důležitá mimo jiné u fenoménů evaginace membrány, tvorby vezikul, mikroklků..
Dva „listy“, které tvoří každou membránu - pamatují na to, že jde o dvojvrstvu - nemají uvnitř stejné složení lipidů; z tohoto důvodu se říká, že distribuce je asymetrická. Tato skutečnost má důležité funkční důsledky.
Specifickým příkladem je složení plazmatické membrány erytrocytů. V těchto krevních buňkách je sfingomyelin a fosfatidylcholin (které vytvářejí membrány s větší relativní tekutostí) obráceny k vnější straně buňky..
Lipidy, které mají tendenci vytvářet tekutější struktury, čelí cytosolu. Za tímto vzorem nenasleduje cholesterol, který je distribuován víceméně homogenně v obou vrstvách..
Funkce membrány každého typu buňky úzce souvisí s její strukturou. Plní však základní funkce.
Biomembrány jsou odpovědné za vymezení buněčného prostředí. Podobně jsou v buňce membránové oddíly..
Například mitochondrie a chloroplasty jsou obklopeny membránami a tyto struktury se účastní biochemických reakcí, ke kterým v těchto organelách dochází..
Membrány regulují průchod materiálů do buňky. Díky této bariéře mohou vstupovat potřebné materiály, ať už pasivně nebo aktivně (s nutností ATP). Nevstupují také nežádoucí nebo toxické materiály.
Membrány udržují iontové složení buňky na odpovídající úrovni prostřednictvím procesů osmózy a difúze. Voda může volně proudit v závislosti na jejím koncentračním gradientu. Soli a metabolity mají specifické transportéry a také regulují buněčné pH.
Díky přítomnosti proteinů a kanálů na povrchu membrány mohou sousední buňky interagovat a vyměňovat si materiály. Tímto způsobem se buňky spojí a vytvoří se tkáně..
A konečně, membrány obsahují značné množství signálních proteinů a umožňují interakci mimo jiné s hormony, neurotransmitery..
Základní složkou membrán jsou fosfolipidy. Tyto molekuly jsou amfipatické, mají polární a apolární zónu. Polární jim umožňuje interakci s vodou, zatímco ocas je hydrofobní uhlíkový řetězec.
Sdružení těchto molekul nastává spontánně ve dvojvrstvě, přičemž hydrofobní ocasy interagují navzájem a hlavy směřují ven..
V malé zvířecí buňce najdeme neuvěřitelně velké množství lipidů, řádově 109 molekuly. Membrány mají tloušťku přibližně 7 nm. Hydrofobní vnitřní jádro, téměř ve všech membránách, zaujímá tloušťku 3 až 4 nm.
Současný model biomembrán je známý jako „tekutá mozaika“, formulovaný v 70. letech výzkumníky Singerem a Nicolsonem. Model navrhuje, aby membrány nebyly tvořeny pouze lipidy, ale také sacharidy a bílkovinami. Termín mozaika označuje tuto směs.
Tvář membrány, která směřuje ven z buňky, se nazývá exoplazmatická tvář. Naproti tomu vnitřní obličej je cytosolický.
Stejná nomenklatura platí pro biomembrány, které tvoří organely, s tou výjimkou, že exoplazmatická tvář v tomto případě směřuje dovnitř buňky, a nikoli ven..
Lipidy, které tvoří membrány, nejsou statické. Ty mají schopnost pohybovat se s určitou mírou volnosti v konkrétních regionech strukturou.
Membrány jsou tvořeny třemi základními typy lipidů: fosfoglyceridy, sfingolipidy a steroidy; všechny jsou amfipatické molekuly. Níže podrobně popíšeme každou skupinu:
První skupina, kterou tvoří fosfoglyceridy, pochází z glycerol-3-fosfátu. Ocas, hydrofobní povahy, je tvořen dvěma řetězci mastných kyselin. Délka řetězů je variabilní: mohou mít od 16 do 18 uhlíků. Mohou mít jednoduché nebo dvojné vazby mezi uhlíky.
Subklasifikace této skupiny je dána typem hlavy, kterou představují. Fosfatidylcholiny jsou nejhojnější a hlava obsahuje cholin. U jiných typů interagují s fosfátovou skupinou různé molekuly, jako je ethanolamin nebo serin.
Další skupinou fosfoglyceridů jsou plaslogeny. Lipidový řetězec je spojen s glycerolem esterovou vazbou; zase existuje uhlíkový řetězec spojený s glycerolem etherovou vazbou. Jsou docela hojné v srdci a mozku.
Sfingolipidy pocházejí ze sfingosinů. Sfingomyelin je hojný sfingolipid. Glykolipidy jsou tvořeny hlavami z cukrů.
Třetí a poslední třída lipidů, které tvoří membrány, jsou steroidy. Jsou to prsteny vyrobené z uhlíků, spojené do čtyřčlenných skupin. Cholesterol je steroid přítomný v membránách a obzvláště hojný u savců a bakterií..
Existují specifické oblasti membrán eukaryotických organismů, kde se koncentruje cholesterol a sfingolipidy. Tyto domény jsou také známé jako vory lipid.
V těchto oblastech také obsahují různé proteiny, jejichž funkcí je buněčná signalizace. Předpokládá se, že lipidové složky modulují proteinové složky v raftech.
V plazmatické membráně je ukotvena řada proteinů. Mohou být integrální, ukotvené k lipidům nebo umístěné na periferii.
Integrály procházejí membránou. Proto musí mít hydrofilní a hydrofobní proteinové domény, aby mohly interagovat se všemi složkami..
V proteinech, které jsou ukotveny k lipidům, je uhlíkový řetězec ukotven v jedné z vrstev membrány. Protein ve skutečnosti nevstupuje do membrány.
Konečně periferní neinteragují přímo s hydrofobní zónou membrány. Spíše mohou být připojeny pomocí integrálního proteinu nebo polárními hlavami. Mohou být umístěny na obou stranách membrány.
Procento proteinů v každé membráně se velmi liší: od 20% v neuronech po 70% v mitochondriální membráně, protože k provádění metabolických reakcí, které se tam vyskytují, potřebuje velké množství proteinových prvků..



Zatím žádné komentáře