The dvojité hnojení je proces, který se vyskytuje v rostlinách, při kterém jedno z gametových jader pylového zrna (mužský gametofyt) oplodňuje ženskou reprodukční buňku (oosféru) a jiné oplodňuje další buňku, což se bude lišit, pokud se jedná o krytosemennou nebo gnetální.
V krytosemenných rostlinách se druhé jádro gametofytu spojuje se dvěma polárními jádry přítomnými v centrální buňce pylového vaku, aby se později vyvinuly v endospermu. U gnetálů naopak druhé jádro mužského gametofytu fúzuje s jádrem ventrálního kanálu a vytváří druhé embryo.
Botanici původně věřili, že dvojité hnojení je výlučným fenoménem krytosemenných rostlin, avšak tento postup byl později popsán také u rostlin skupiny gnetal..
Rejstřík článků
Botanik ruského původu Sergej Gavrilovič Nawashin pracující s krytosemennými druhy Lilium martagon Y Fritillaria tenella byl první, kdo sledoval proces dvojitého oplodnění. Tento jev později demonstroval slavný polsko-německý botanik Eduard Strasburger.
Po tomto objevu hlásili různí botanici abnormální události dvojitého oplodnění v různých skupinách gymnospermů (Thuja, Abies, Pseudotsuga, mimo jiné). Produkty druhého oplodnění by mohly degenerovat, produkovat volná jádra nebo také způsobit další embrya.
Později se ukázalo, že dvojité oplodnění bylo normální událostí u rostlin skupiny gnetal, ale u nich, na rozdíl od krytosemenných rostlin, druhé oplodnění vždy produkuje další embrya a ne endospermy..
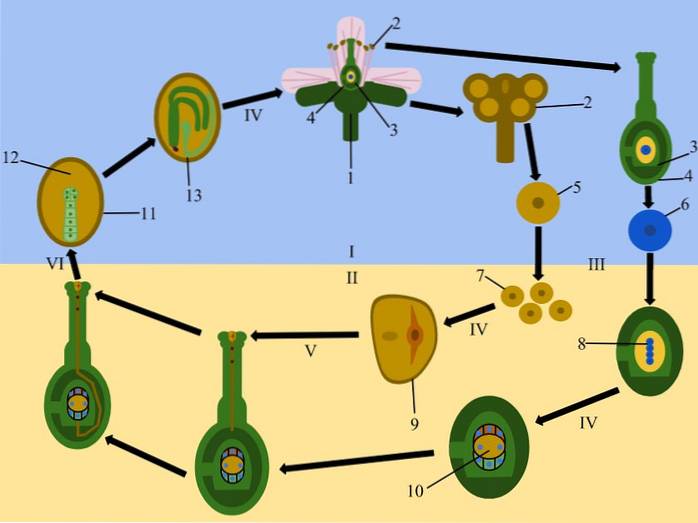
Ve většině krytosemenných rostlin produkuje prekurzorová buňka megasporů (megasporocyt) čtyři haploidní megaspory meiózou, z nichž pouze jeden se vyvíjí a produkuje megagametofyt, zatímco ostatní degenerují.
Megagametofyt produkuje osm jader, z nichž dvě (polární jádra) migrují směrem k centrální zóně megagametofytu nebo embryonálního vaku, čímž vznikají dvoujaderné buňky..
Zbytek jader je uspořádán do skupin po třech na periferii, z nichž jedna bude tvořit ovocell, dvě sousední budou tvořit synergie, zatímco zbývající tři umístěné na opačném konci budou tvořit antipody..
Samčí gametofyt (pylové zrno) produkuje tři jádra; dvě spermatické a jedna vegetativní. Když pylové zrno přijde do styku se stigmatem, vyklíčí a vytvoří pylovou trubici, která roste stylem a přitahuje ji látky produkované synergenty..
Dvě jádra spermií poté migrují pylovou trubicí a provádějí dvojité oplodnění. Jedno z jader spermií se spojí s jádrem vaječné buňky a vytvoří zygotu, která povede ke vzniku embrya, zatímco druhé se spojí se dvěma jádry centrální buňky, která způsobí vznik endospermu.
Endosperm je triploidní tkáň vytvořená fúzí centrální buňky embryonálního vaku (binucleate) s jedním ze dvou spermií jádra pylového zrna. Buňky endospermu jsou bohaté na škrobové granule zabudované do proteinové matrice a jejich funkcí je dodávat výživný materiál do vyvíjejícího se embrya.
Mezi vědci neexistuje shoda ohledně evolučního původu endospermu v krytosemenných rostlinách. Někteří autoři tvrdí, že se jedná o extra embryo, jehož vývoj byl transformován na výživnou tkáň ve prospěch druhého embrya.
Jiní autoři tvrdí, že endosperm není produktem sexuální reprodukce, ale je vegetativní fází vývoje embryonálního vaku, jak se vyskytuje u gymnospermů. Obě hypotézy mají své kritiky a obhájce.
Jediné rostliny gymnospermu, u nichž je prokázáno dvojité hnojení, patří do rodů Gnetum Y Ephedra (Gnatales). U žádného z nich však endosperm nepochází z tohoto dvojitého oplodnění..
U nejméně dvou druhů Ephedra ženské gamety se tvoří v archegonii v monosporických gametofytech. Spermatická trubice obsahuje čtyři jádra, z nichž dvě jsou reprodukční.
Tato jádra spermií se uvolňují v megagametofytu a jedno z nich se spojí s jádrem vaječné buňky, zatímco druhé se spojí s jádrem ventrálního kanálu..
Výsledkem jsou dva zygoti, z nichž se vyvinou geneticky identická životaschopná embrya, protože obě ženská jádra byla vytvořena v monosporickém gametofytu, zatímco dvě jádra spermií ze stejné pylové trubice jsou také geneticky identická..
V každé samičí gametofytu se mohou tvořit dvě nebo více archegonií, což má za následek více událostí dvojitého oplodnění, které se v gametofytu vyskytují současně..
Dvojité hnojení v Gnetum představuje důležité rozdíly ve srovnání s dvojitým oplodněním, ke kterému dochází v Ephedra. Na Gnetum, na rozdíl od v Ephedra, ženské gametofyty jsou tetrasporické a ne monosporické.
Další rozdíl je v tom Gnetum netvoří se ani archegonia, ani předurčené ovocell. Kvůli ztrátě oocelů je oplodnitelné mnoho ženských gametofytických jader; tímto způsobem mohou dvě jádra spermií z pylové trubice oplodnit kterákoli dvě ženská jádra.
Jako v Ephedra, proces dvojitého hnojení v Gnetum bude produkovat dva životaschopné zygoty, ale v tomto nebudou zygoty geneticky identické kvůli tetrasporické povaze ženského gametofytu. Na Gnetum může dojít také k několika událostem dvojitého oplodnění, pokud je dostatek pylových zrn.

Objev dvojitého oplodnění v Gnetum Y Ephedra podporuje evoluční hypotézu, podle které tento proces vznikl u společného předka Gnetales a krytosemenných rostlin, pro něž by to byla synapomorfie (sdílený odvozený znak), která by jim umožnila seskupit je do skupiny antofytů (monofyletických).
Synapomorphies jsou odvozené znaky, které jsou sdíleny dvěma nebo více druhy nebo taxony, a proto mohou vykazovat určitý stupeň příbuznosti. Rodový charakter (plesiomorfní) by v takovém případě bylo jednoduché oplodnění.
Tímto způsobem by Gnetales mohli být bazální skupinou v rámci anthofytů, ve kterých dvojité oplodnění vede ke dvěma životaschopným zygotům, zatímco vzhled endospermu jako produktu dvojitého oplodnění by byl jedinečnou synapomorfií v krytosemenných rostlinách..

Zatím žádné komentáře