The zákon o ekologickém desátku, ekologický zákon nebo 10% zvyšuje způsob, jakým energie putuje ve své derivaci různými trofickými úrovněmi. Často se také tvrdí, že tento zákon je jednoduše přímým důsledkem druhého zákona termodynamiky.
Ekologická energie je součástí ekologie, která se zabývá kvantifikací vztahů, které jsme načrtli výše. Má se za to, že Raymond Lindemann (konkrétně ve své klíčové práci z roku 1942) byl tím, kdo založil základy této oblasti studia.
Jeho práce se zaměřila na koncepty potravinového řetězce a webu a na kvantifikaci účinnosti při přenosu energie mezi různými trofickými úrovněmi..
Lindemann vychází z dopadajícího slunečního záření nebo energie, kterou komunita přijímá, prostřednictvím zachycení prováděného rostlinami pomocí fotosyntézy a pokračuje v monitorování tohoto zachycení a jeho následného použití býložravci (primární konzumenti), poté masožravci (sekundární konzumenti) a nakonec rozkladače.
Rejstřík článků
Po Lindemannově průkopnické práci se předpokládalo, že trofická účinnost přenosu bude kolem 10%; ve skutečnosti někteří ekologové odkazovali na 10% zákon. Od té doby však došlo k několika zmatkům ohledně tohoto problému..
Určitě neexistuje žádný přírodní zákon, který by vedl k tomu, že přesně jedna desetina energie vstupující do jedné trofické úrovně bude přenesena na další..
Například kompilace trofických studií (v mořském a sladkovodním prostředí) odhalila, že účinnost přenosu podle trofické úrovně se pohybovala mezi přibližně 2 a 24%, ačkoli průměr byl 10,13%..
Obecně platí, že platí pro vodní i suchozemské systémy, lze říci, že sekundární produktivita býložravců se obvykle nachází přibližně, řádově pod primární produktivitou, na které je založena..
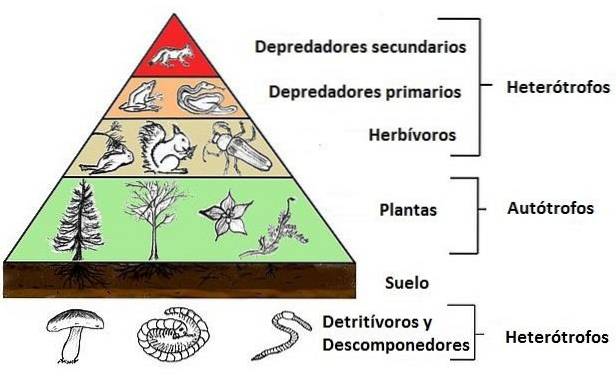
Jedná se často o konzistentní vztah, který se udržuje ve všech pástových systémech a který má tendenci se stát strukturami pyramidového typu, ve kterých základnu poskytují rostliny a na této základně je založen menší, primárních spotřebitelů, na nichž je jiný (i menší) sekundární spotřebitelé sedí.
Všechny živé věci vyžadují hmotu a energii; hmota pro stavbu jejich těl a energii k provádění jejich životních funkcí. Tento požadavek není omezen na jednotlivý organismus, ale je rozšířen na vyšší úrovně biologické organizace, které tito jedinci mohou vyhovovat..
Tyto úrovně organizace jsou:
V ekosystému vytváří komunita a životní prostředí toky energie a hmoty.
Organismy ekosystému jsou seskupeny podle „role“ nebo „funkce“, kterou plní v potravním nebo trofickém řetězci; takto mluvíme o trofických úrovních producentů, spotřebitelů a rozkladačů.
Každá z těchto trofických úrovní zase interaguje s fyzikálně-chemickým prostředím, které poskytuje podmínky pro život a současně působí jako zdroj a jímka energie a hmoty..
Nejprve musíme definovat primární produktivitu, což je rychlost produkce biomasy na jednotku plochy..
Obvykle se vyjadřuje v jednotkách energie (jouly na metr čtvereční za den) nebo v jednotkách suché organické hmoty (kilogramy na hektar a rok) nebo jako uhlík (hmotnost uhlíku v kg na metr čtvereční za rok).
Obecně, když mluvíme o veškeré energii fixované fotosyntézou, obvykle to nazýváme hrubá primární produktivita (PPG).
Z toho část je vynaložena na dýchání stejných autotrofů (RA) a je ztracena ve formě tepla. Čistá primární produkce (PPN) se získá odečtením tohoto množství od PPG (PPN = PPG-RA).
Tato čistá primární produkce (PPN) je to, co je nakonec k dispozici ke spotřebě heterotrofy (jedná se o bakterie, houby a ostatní zvířata, která známe).
Sekundární produktivita (PS) je definována jako rychlost produkce nové biomasy heterotrofními organismy. Na rozdíl od rostlin, heterotrofních bakterií, hub a zvířat nemohou z jednoduchých molekul vytvořit složité a energeticky bohaté sloučeniny, které potřebují..
Vždy získávají svoji hmotu a energii z rostlin, což mohou dělat přímo konzumací rostlinného materiálu nebo nepřímo krmením jinými heterotrofy..
Je to tak, že rostliny nebo fotosyntetické organismy obecně (nazývané také producenti) tvoří první trofickou úroveň v komunitě; primární konzumenti (ti, kteří se živí producenty) tvoří druhou trofickou úroveň a sekundární konzumenti (nazývaní také masožravci) tvoří třetí úroveň.
Podíly čisté primární produkce, které tečou po každé z možných energetických drah, v konečném důsledku závisí na účinnosti přenosu, tj. Na způsobu, jakým je energie využívána a předávána z jedné úrovně do druhé..
Existují tři kategorie účinnosti přenosu energie a s těmito dobře definovanými možnostmi můžeme předvídat vzorec toku energie na trofických úrovních. Jedná se o tyto kategorie: účinnost spotřeby (EC), účinnost asimilace (EA) a účinnost výroby (EP).
Pojďme nyní definovat tyto tři zmíněné kategorie.
Matematicky můžeme definovat účinnost spotřeby (EC) následovně:
EC =Ján/Pn-1 × 100
Kde vidíme, že CE je procento z celkové dostupné produktivity (Pn-1) který je účinně pohlcován horním sousedícím trofickým oddílem (Ján).
Například pro primární spotřebitele v systému pastvy je EC procento (vyjádřeno v jednotkách energie a na jednotku času) PPN, které je spotřebováno býložravci.
Pokud bychom mluvili o sekundárních spotřebitelích, pak by to odpovídalo procentu produktivity býložravců, které konzumují masožravci. Zbytek zemře nezjištěný a vstoupí do řetězce rozpadu.
Na druhou stranu je účinnost asimilace vyjádřena takto:
EA =NAn/Ján × 100
Opět odkazujeme na procento, ale tentokrát na část energie pocházející z potravy a spotřebovanou spotřebitelem v trofické komoře (Ján) a to je asimilováno trávicím systémem (NAn).
Tato energie bude ta, která je k dispozici pro růst a pro provádění práce. Zbytek (část, která není asimilována) se ztratí spolu s výkaly a poté vstoupí do trofické úrovně rozkladačů.
Nakonec je účinnost výroby (EP) vyjádřena jako:
EP = Pn/NAn × 100
což je také procento, ale v tomto případě odkazujeme na asimilovanou energii (NAn), který je nakonec začleněn do nové biomasy (Pn). Veškerý zbytek energie, který není asimilován, se během dýchání ztrácí ve formě tepla.
Produkty, jako jsou sekrece a / nebo exkrece (bohaté na energii), které se účastnily metabolických procesů, lze považovat za produkci, Pn, a jsou k dispozici, jako mrtvoly, rozkladačům.
Po definování těchto tří důležitých kategorií si nyní můžeme položit otázku o „globální efektivitě přenosu“ z jedné trofické úrovně na druhou, která je jednoduše dána součinem výše zmíněné efektivity (EC x EA x EP).
Řečeno hovorově, můžeme říci, že účinnost hladiny je dána tím, co lze účinně pohltit, což se pak asimiluje a nakonec se začlení do nové biomasy.
Produktivita býložravců je vždy nižší než produktivita rostlin, kterými se živí. Pak bychom si mohli položit otázku: Kam jde zbytečná energie?
Abychom mohli odpovědět na tuto otázku, musíme upozornit na následující fakta:
To se děje ze dvou základních důvodů: Zaprvé kvůli tomu, že neexistuje 100% efektivní proces přeměny energie. To znamená, že při přeměně vždy dochází ke ztrátě ve formě tepla, což je v dokonalé shodě s druhým zákonem termodynamiky..
Zadruhé, protože zvířata musí dělat práci, která vyžaduje energetický výdej, což má za následek nové ztráty v podobě tepla..
Tyto vzorce se vyskytují na všech trofických úrovních a jak předpovídá druhý zákon termodynamiky, část energie, kterou se člověk snaží přenést z jedné úrovně do druhé, je vždy rozptýlena ve formě nepoužitelného tepla..


Zatím žádné komentáře