
The endoplazmatické retikulum je to membránová buněčná organela přítomná ve všech eukaryotických buňkách. Tento složitý systém zabírá přibližně více než polovinu membrán v běžné zvířecí buňce. Membrány pokračují, dokud se nesetkají s jadernou membránou a tvoří souvislý prvek.
Tato struktura je distribuována v buněčné cytoplazmě ve formě labyrintu. Jedná se o druh sítě tubulů navzájem propojených vakovitými strukturami. V endoplazmatickém retikulu dochází k biosyntéze proteinů a lipidů. Téměř všechny proteiny, které musí být přeneseny ven z buňky, nejprve projdou retikulem.
Membrána retikula není odpovědná pouze za oddělení vnitřku této organely od cytoplazmatického prostoru a zprostředkování transportu molekul mezi těmito buněčnými oddíly; Podílí se také na syntéze lipidů, které budou součástí plazmatické membrány buňky a membrán ostatních organel..
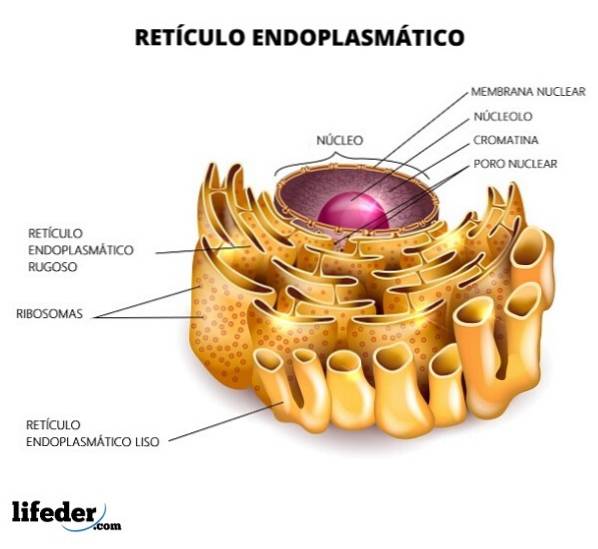
Retikulum je rozděleno na hladké a drsné, v závislosti na přítomnosti nebo nepřítomnosti ribozomů v jeho membránách. Drsné endoplazmatické retikulum má ribozomy připojené k membráně (přítomnost ribozomů jí dodává „drsný“ vzhled) a tvar tubulů je mírně rovný.
Hladké endoplazmatické retikulum postrádá ribozomy a tvar struktury je mnohem nepravidelnější. Funkce hrubého endoplazmatického retikula je zaměřena hlavně na zpracování proteinů. Hladký je naopak zodpovědný za metabolismus lipidů.
Rejstřík článků
Endoplazmatické retikulum je membránová síť přítomná ve všech eukaryotických buňkách. Skládá se ze vakul nebo cisteren a tubulárních struktur, které tvoří kontinuum s membránou jádra a jsou distribuovány po celé buňce.
Lumen retikula se kromě oxidačního prostředí vyznačuje vysokou koncentrací iontů vápníku. Obě vlastnosti mu umožňují plnit své funkce.
Endoplazmatické retikulum je považováno za největší organelu přítomnou v buňkách. Objem buňky v tomto oddělení pokrývá přibližně 10% vnitřku buňky.
Drsné endoplazmatické retikulum má na povrchu vysokou hustotu ribozomů. Je to oblast, kde dochází ke všem procesům souvisejícím se syntézou a modifikací bílkovin. Jeho vzhled je hlavně trubkovitý.
Hladké endoplazmatické retikulum nemá ribozomy. Je hojný v buněčných typech, které mají aktivní metabolismus při syntéze lipidů; například v buňkách varlat a vaječníků, které jsou buňkami produkujícími steroidy.
Podobně hladké endoplazmatické retikulum se nachází v poměrně vysokém podílu v jaterních buňkách (hepatocytech). V této oblasti dochází k produkci lipoproteinů..
Ve srovnání s hrubým endoplazmatickým retikulem je jeho struktura komplikovanější. Hojnost hladkého versus hrubého retikula závisí především na typu buňky a její funkci..
Fyzická architektura endoplazmatického retikula je spojitý systém membrán tvořených vzájemně propojenými vaky a tubuly. Tyto membrány sahají až k jádru a tvoří jediný lumen.
Mříž je postavena několika doménami. Distribuce je spojena s jinými organelami, různými proteiny a složkami cytoskeletu. Tyto interakce jsou dynamické.
Strukturálně se endoplazmatické retikulum skládá z jaderného obalu a periferního endoplazmatického retikula, tvořeného tubuly a vaky. Každá struktura souvisí s konkrétní funkcí.
Jaderný obal, stejně jako všechny biologické membrány, je tvořen lipidovou dvojvrstvou. Tím je ohraničen interiér sdílený s periferním retikulem.
Vaky, které tvoří endoplazmatické retikulum, jsou ploché a často skládané. Obsahují zakřivené oblasti na okrajích membrán. Trubková síť není statická entita; může růst a restrukturalizovat.
Systém vaků a tubulů je přítomen ve všech eukaryotických buňkách. Liší se však tvarem a strukturou v závislosti na typu buňky..
Retikulum buněk s důležitými funkcemi při syntéze bílkovin je složeno převážně z vaků, zatímco buňky nejvíce související se syntézou lipidů a signalizací vápníku jsou složeny z většího počtu tubulů..
Příkladem buněk s vysokým počtem vaků jsou sekreční buňky pankreatu a buňky B. Naproti tomu svalové buňky a jaterní buňky mají síť prominentních tubulů..
Endoplazmatické retikulum je zapojeno do řady procesů, včetně syntézy proteinů, přenosu a skládání a modifikací, jako je tvorba disulfidové vazby, glykosylace a přidání glykolipidů. Kromě toho se podílí na biosyntéze membránových lipidů.
Nedávné studie spojily retikulum s odpověďmi na buněčný stres a mohou dokonce vyvolat procesy apoptózy, i když mechanismy nebyly zcela objasněny. Všechny tyto procesy jsou podrobně popsány níže:
Endoplazmatické retikulum je úzce spjato s obchodováním s bílkovinami; konkrétně na proteiny, které musí být zaslány do zahraničí, na Golgiho aparát, na lysosomy, na plazmatickou membránu a logicky na ty, které patří do stejného endoplazmatického retikula.
Endoplazmatické retikulum je buněčné chování podílející se na syntéze proteinů, které musí být přenášeno mimo buňku. Tato funkce byla objasněna skupinou vědců v 60. letech, která studovala buňky pankreatu, jejichž funkcí je vylučovat trávicí enzymy..
Tato skupina, vedená Georgem Paladeem, dokázala značit proteiny pomocí radioaktivních aminokyselin. Tímto způsobem bylo možné vystopovat a lokalizovat proteiny technikou zvanou autoradiografie..
Radioaktivně značené proteiny lze vysledovat do endoplazmatického retikula. Tento výsledek naznačuje, že retikulum se účastní syntézy proteinů, jejichž konečným cílem je sekrece..
Později se proteiny přesunou do Golgiho aparátu, kde jsou „zabaleny“ do vezikul, jejichž obsah bude vylučován..
K procesu sekrece dochází, protože membrána vezikul může fúzovat s plazmatickou membránou buňky (obě mají lipidovou povahu). Tímto způsobem může být obsah uvolněn ven z buňky..
Jinými slovy, vylučované proteiny (a také proteiny zaměřené na lysozomy a plazmatickou membránu) musí následovat specifickou cestu, která zahrnuje drsné endoplazmatické retikulum, Golgiho aparát, sekreční váčky a nakonec zevnějšek buňky..
Proteiny, které jsou určeny k zabudování do nějaké biomembrány (plazmatická membrána, Golgiho membrána, lysozom nebo retikulum), jsou nejprve vloženy do membrány retikula a nejsou uvolňovány okamžitě do lumenu. Musí se řídit stejnou cestou pro sekreční proteiny.
Tyto proteiny mohou být umístěny uvnitř membrán hydrofobním sektorem. Tato oblast má řadu 20 až 25 hydrobofických aminokyselin, které mohou interagovat s uhlíkovými řetězci fosfolipidů. Způsob, jakým se tyto proteiny vkládají, je však variabilní..
Mnoho proteinů prochází membránou pouze jednou, zatímco jiné opakovaně. Podobně to může být v některých případech konec karboxylového konce nebo amino konce.
Orientace uvedeného proteinu je stanovena, zatímco peptid roste a je přenášen do endoplazmatického retikula. Všechny proteinové domény, které směřují k lumenu retikula, budou nalezeny na vnější straně buňky v jejich konečné poloze..
Molekuly proteinové povahy mají trojrozměrnou konformaci nezbytnou k provádění všech svých funkcí..
DNA (kyselina deoxyribonukleová), procesem nazývaným transkripce, předává své informace molekule RNA (kyselina ribonukleová). RNA poté prochází do proteinů procesem translace. Peptidy se přenesou do retikula, když probíhá proces translace.
Tyto řetězce aminokyselin jsou v retikulu uspořádány trojrozměrně pomocí proteinů nazývaných chaperony: protein rodiny Hsp70 (proteiny tepelného šoku nebo proteiny tepelného šoku pro jeho zkratku v angličtině; číslo 70 označuje jeho atomovou hmotnost 70 KDa) zvanou BiP.
BiP protein se může vázat na polypeptidový řetězec a zprostředkovat jeho skládání. Podobně se podílí na sestavení různých podjednotek, které tvoří kvartérní strukturu proteinů..
Proteiny, které nebyly správně složeny, jsou zadržovány retikulem a zůstávají vázány na BiP nebo se degradují.
Když je buňka vystavena stresovým podmínkám, retikulum na ni reaguje a v důsledku toho nedochází ke správnému skládání proteinů. Buňka se může obrátit na jiné systémy a produkovat bílkoviny, které udržují homeostázu retikula.
Disulfidový můstek je kovalentní vazba mezi sulfhydrylovými skupinami, které jsou součástí aminokyselinové struktury cysteinu. Tato interakce je zásadní pro fungování určitých proteinů; podobně definuje strukturu proteinů, které je prezentují.
Tyto vazby nelze vytvořit v jiných buněčných kompartmentech (například v cytosolu), protože nemají oxidační prostředí, které upřednostňuje jeho tvorbu..
Na tvorbě (a lámání) těchto vazeb se podílí enzym: protein disulfid izomeráza.
V retikulu probíhá proces glykosylace u konkrétních zbytků asparaginu. Stejně jako skládání proteinů dochází k glykosylaci, když běží proces translace.
Oligosacharidové jednotky jsou tvořeny čtrnácti zbytky cukru. Přenášejí se na asparagin pomocí enzymu nazývaného oligosacaryltransferáza, který se nachází v membráně.
Zatímco je protein v retikulu, jsou odstraněny tři zbytky glukózy a jeden zbytek manózy. Tyto proteiny jsou odváděny do Golgiho aparátu k dalšímu zpracování..
Na druhou stranu určité proteiny nejsou ukotveny k plazmatické membráně částí hydrofobních peptidů. Naproti tomu jsou připojeny k určitým glykolipidům, které fungují jako kotevní systém, a nazývají se glykosylfosfatidylinositol (zkráceně GPI)..
Tento systém je shromážděn v membráně retikula a zahrnuje vazbu GPI na koncový uhlík proteinu..
Endoplazmatické retikulum hraje klíčovou roli v biosyntéze lipidů; konkrétně hladké endoplazmatické retikulum. Lipidy jsou nepostradatelnou součástí plazmatických membrán buněk.
Lipidy jsou vysoce hydrofobní molekuly, takže je nelze syntetizovat ve vodném prostředí. Proto k jeho syntéze dochází ve spojení s již existujícími membránovými složkami. Transport těchto lipidů nastává ve váčcích nebo transportními proteiny.
Membrány eukaryotických buněk jsou tvořeny třemi typy lipidů: fosfolipidy, glykolipidy a cholesterol..
Fosfolipidy jsou odvozeny od glycerolu a jsou nejdůležitější strukturní složkou. Ty jsou syntetizovány v oblasti membrány retikula, která ukazuje na cytosolický obličej. Na procesu se podílejí různé enzymy.
Membrána roste integrací nových lipidů. Díky existenci enzymové flipázy může růst nastat v obou polovinách membrány. Tento enzym je zodpovědný za přenos lipidů z jedné strany dvojvrstvy na druhou..
Procesy syntézy cholesterolu a ceramidů se také vyskytují v retikulu. Ten cestuje do Golgiho aparátu za účelem výroby glykolipidů nebo sfingomyelinu..
Molekula vápníku se účastní jako signalizátor různých procesů, ať už jde o fúzi nebo asociaci proteinů s jinými proteiny nebo s nukleovými kyselinami..
Vnitřek endoplazmatického retikula má koncentrace vápníku 100-800 uM. Vápníkové kanály a receptory, které uvolňují vápník, se nacházejí v retikulu. Uvolňování vápníku nastává, když je fosfolipáza C stimulována aktivací receptorů spojených s G-proteinem (GPCR).
Dále dochází k eliminaci fosfatidylinositol 4,5-bisfosfátu v diacylglycerolu a inositol trifosfátu; druhý je zodpovědný za uvolňování vápníku.
Svalové buňky mají endoplazmatické retikulum specializované na sekvestraci iontů vápníku, které se nazývá sarkoplazmatické retikulum. Podílí se na procesech svalové kontrakce a relaxace.



Zatím žádné komentáře