
The teorie skládání membrán navrhuje, aby organelové membrány pocházely z expanze a invaginace plazmatické membrány. J.D Robertson, průkopník v elektronové mikroskopii, v roce 1962 zaznamenal, že několik intracelulárních těl má strukturu, která je vizuálně identická s plazmatickou membránou..
Myšlenka struktury, která vymezuje buňky, vznikla bezprostředně po vzniku pojmu „buňka“, pro který bylo provedeno několik studií k objasnění charakteristik uvedené struktury..
Rejstřík článků
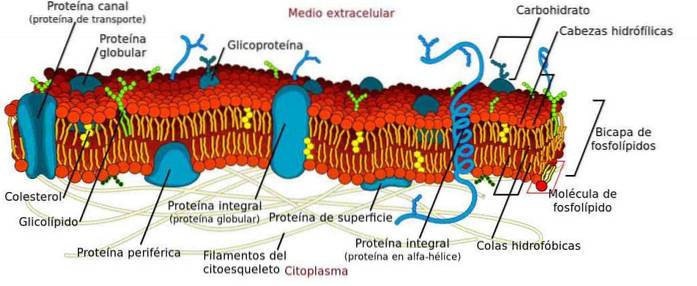
Plazmatická membrána je struktura tvořená dvojitou vrstvou fosfolipidů uspořádaných takovým způsobem, že polární skupiny jsou orientovány směrem k cytosolu a extracelulárnímu médiu, zatímco nepolární skupiny jsou uspořádány směrem dovnitř membrány..
Jeho hlavní funkcí je definovat buňky, eukaryotické i prokaryotické, protože fyzicky odděluje cytoplazmu od extracelulárního prostředí..
Navzdory své strukturální funkci je dobře známo, že membrána není statická, ale spíše elastická a dynamická bariéra, kde dochází k velkému počtu základních procesů pro buňku..
Některé procesy, které probíhají v membráně, jsou ukotvení cytoskeletu, transport molekul, signalizace a spojení s jinými buňkami za vzniku tkání. Kromě toho má široká škála organel také membránu, ve které probíhají další velmi důležité procesy..
Dlouho předtím, než Robertson v roce 1962 navrhl teorii skládání membrán, byly nejprve provedeny studie, aby se zjistilo, jak tato struktura vypadá. Při absenci elektronového mikroskopu převládaly elektrofyziologické studie, mezi které patří:
Overton poznamenal, že lipidy procházely buněčnou membránou snadněji než molekuly jiné povahy, a proto vyvodil, že membrána musí být z větší části složena z lipidů.
J. Bernstein představil svou hypotézu, která zmínila, že buňky sestávaly z roztoku s volnými ionty ohraničenými tenkou vrstvou nepropustnou pro uvedené nabité molekuly..
Fricke měřil schopnost membrány erytrocytů ukládat náboje (kapacitu) a určoval, že tato hodnota byla 0,81 µF / cmdva.
Později bylo zjištěno, že membrány jiných typů buněk měly podobné hodnoty kapacity, proto by membrána měla mít jednotnou strukturu.
Gorter a Grendel měřili pomocí mikroskopu plochu savčích erytrocytů. Poté extrahovali lipidy ze známého počtu tohoto typu buněk a změřili plochu, kterou zabírali..
Výsledkem byl poměr buněk: membrán 1: 2. To znamenalo, že buněčná membrána měla dvojitou strukturu, což dalo vzniknout pojmu „lipidová dvojvrstva“.
Studie před rokem 1935 naznačovaly přítomnost proteinů v membráně, což vedlo Danielliho a Davsona k návrhu Sandwichova modelu nebo modelu Protein-Lipid-Protein..
Podle tohoto modelu se plazmatická membrána skládá ze dvou vrstev fosfolipidů, které se nacházejí mezi dvěma vrstvami proteinů, které jsou s membránou spojeny elektrostatickými interakcemi..
V roce 1959 shromáždil J. David Robertson díky elektronové mikroskopii dostatek důkazů, aby potvrdil a doplnil modely navržené Gorterem a Grendelem (1925) a Danielli a Davsonem (1935) a navrhl model „Unitary Membrane“..
Tento model si zachovává charakteristiku modelu, který navrhli Danielli a Davson lipidové dvojvrstvy, se změnou proteinové vrstvy, která je v tomto případě asymetrická a diskontinuální. .
Příchod elektronové mikroskopie umožnil mít docela jasnou představu o tom, jak byla vytvořena plazmatická membrána.
Tato skutečnost však byla doprovázena vizualizací několika intracytoplazmatických membrán, které tvořily intracelulární kompartmenty, což vedlo Robertsona v roce 1962 k návrhu „Teorie skládání membrány“.
Teorie skládání membrány spočívá v tom, že plazmatická membrána zvětšila svůj povrch a invaginovala za vzniku intracytoplazmatických membrán, které obklopovaly molekuly, které byly v cytosolu, čímž vznikly organely.
Podle této teorie mohl takto vzniknout jaderný obal, endoplazmatické retikulum, Golgiho aparát, lysozomy a vakuoly..
Kontinuita, která existuje mezi plazmatickou membránou a prvními třemi organelami zmíněnými výše, byla potvrzena studiemi elektronové mikroskopie u různých typů buněk..
Robertson však ve své teorii také navrhl, že vezikulární organely, jako jsou lysosomy a vakuoly, také vznikly invaginacemi, které se následně oddělily od membrány..
Vzhledem k charakteristikám teorie skládání membrán se považuje za rozšíření modelu jednotkové membrány, který sám navrhl v roce 1959.
Mikrografy pořízené Robertsonem ukazují, že všechny tyto membrány jsou stejné, a proto by měly mít poměrně podobné složení..
Specializace organel však značně modifikuje složení membrán, což snižuje jejich společné vlastnosti na biochemické a molekulární úrovni..
Stejným způsobem je zachována skutečnost, že hlavní funkcí membrán je sloužit jako stabilní bariéra pro vodná média..
Díky všem testům provedeným v letech 1895 až 1965, zejména mikroskopickým studiím provedeným J.D. Robertsone, byl zdůrazněn význam buněčných membrán.
Od jeho jednotného modelu začala být zdůrazňována základní role, kterou hraje membrána ve struktuře a funkci buněk, až do té míry, že studium této struktury je považováno za základní problém současné biologie..
S ohledem na příspěvek teorie skládání membrán to však v současné době není přijato. V té době to však vedlo k tomu, že se více odborníků v této oblasti pokoušelo objasnit původ nejen buněčných membrán, ale také původ samotné eukaryotické buňky, jak to udělal Lynn Margulis v roce 1967 při zvyšování endosymbiotické teorie..



Zatím žádné komentáře