The topoizomerázy Jedná se o typ izomerázových enzymů, které modifikují topologii deoxyribonukleové kyseliny (DNA) a generují její odvíjení i navíjení a navíjení.
Tyto enzymy mají specifickou roli při zmírňování torzního stresu v DNA, takže mohou nastat důležité procesy, jako je replikace, transkripce DNA na messenger ribonukleovou kyselinu (mRNA) a rekombinace DNA..
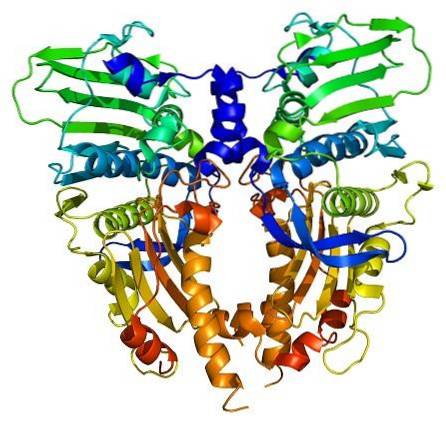
Enzymy topoizomerázy jsou přítomny v eukaryotických i prokaryotických buňkách. Jeho existenci předpověděli vědci Watson a Crick při hodnocení omezení, která struktura DNA představovala, aby umožnila přístup k jejím informacím (uloženým v jeho nukleotidové sekvenci).
Abychom porozuměli funkcím topoizomeráz, je třeba mít za to, že DNA má stabilní strukturu dvoušroubovice, jejíž vlákna jsou navinuta jedna přes druhou..
Tyto lineární řetězce jsou tvořeny 2-deoxyribózou spojenou 5'-3 'fosfodiesterovými vazbami a dusíkatými bázemi uvnitř nich, jako příčky točitého schodiště..
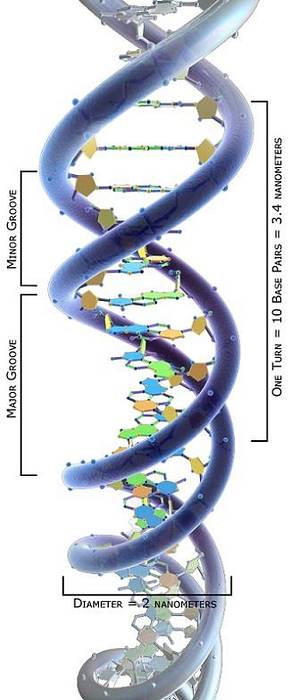
Topologická studie molekul DNA ukázala, že mohou předpokládat různé konformace závislé na jejich torzním namáhání: od uvolněného stavu po různé navíjecí stavy, které umožňují jejich zhutnění..
Molekuly DNA s různými konformacemi se nazývají topoizomery. Můžeme tedy dojít k závěru, že topoizomerázy I a II mohou zvyšovat nebo snižovat torzní napětí molekul DNA a vytvářet jejich různé topoizomery..
Mezi možnými topoizomery DNA je nejběžnější konformací supercoil, který je velmi kompaktní. Dvojitá šroubovice DNA však musí být také odvíjena topoizomerázami během různých molekulárních procesů..
Rejstřík článků
Některé topoizomerázy mohou uvolňovat pouze negativní supercoily DNA, nebo obě supercoily DNA: pozitivní i negativní.
Pokud se kruhová dvouvláknová DNA odvíjí na své podélné ose a dojde k otočení levou rukou (ve směru hodinových ručiček), říká se, že je negativně navinutá. Pokud je otočení ve směru hodinových ručiček (proti směru hodinových ručiček), je pozitivně převinuto.

Topoizomerázy mohou v zásadě:
-Usnadněte průchod řetězce DNA řezem v opačném řetězci (topoizomeráza typu I).
-Usnadněte průchod úplné dvojité šroubovice samotným štěpením nebo štěpením do jiné dvojité šroubovice (topoizomeráza typu II).
Stručně řečeno, topoizomerázy působí štěpením fosfodiesterových vazeb v jednom nebo obou řetězcích, které tvoří DNA. Poté upraví stav navíjení pramenů dvojité šroubovice (topoizomeráza I) nebo dvou dvojitých šroubovic (topoizomeráza II), aby konečně znovu spojily nebo spojily rozštěpené konce.
Ačkoli topoizomeráza I je enzym, který vykazuje vyšší aktivitu během S fáze (syntéza DNA), nepovažuje se za závislou na fázi buněčného cyklu.
Zatímco aktivita topoizomerázy II je nejaktivnější během logaritmické fáze růstu buněk a v buňkách rychle rostoucích nádorů.
Změna genů kódujících topoizomerázy je pro buňky smrtelná, což dokazuje důležitost těchto enzymů. Mezi procesy, kterých se topoizomerázy účastní, patří:
Topoizomerázy usnadňují kompaktní ukládání genetické informace, protože vytvářejí navíjení a nadzávitky DNA, což umožňuje najít velké množství informací v relativně malém objemu.
Bez topoizomeráz a jejich jedinečných charakteristik by byl přístup k informacím uloženým v DNA nemožný. To je způsobeno skutečností, že topoizomerázy periodicky uvolňují torzní napětí, které je generováno v dvojité šroubovici DNA během jejího odvíjení v procesech replikace, transkripce a rekombinace..
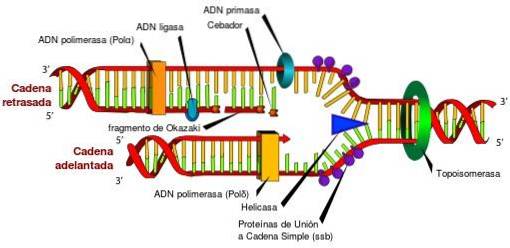
Pokud se torzní napětí generované během těchto procesů neuvolní, může dojít k defektní genové expresi, přerušení kruhové DNA nebo chromozomu, dokonce k produkci buněčné smrti.
Konformační změny (v trojrozměrné struktuře) molekuly DNA vystavují vnější oblasti, které mohou interagovat s proteiny vázajícími DNA. Tyto proteiny mají regulační funkci genové exprese (pozitivní nebo negativní).
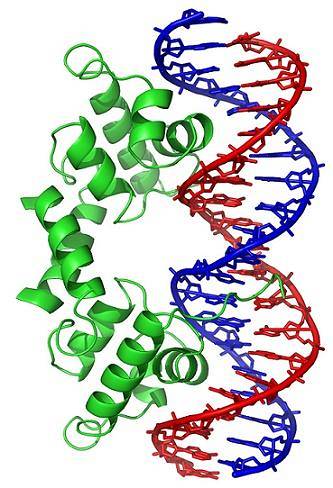
Stav navíjení DNA, generovaný působením topoizomeráz, tedy ovlivňuje regulaci genové exprese.
Topoizomeráza II je nezbytná pro shromažďování chromatidů, kondenzaci a dekondenzaci chromozomů a segregaci dceřiných molekul DNA během mitózy..
Tento enzym je také strukturním proteinem a jednou z hlavních složek matice buněčného jádra během mezifáze..
Existují dva hlavní typy topoizomeráz podle toho, zda jsou schopny štěpit jeden nebo dva řetězce DNA.
Topoizomerázy typu I jsou monomery, které zmírňují negativní a pozitivní supercoily, které jsou vytvářeny pohybem vlásenky během transkripce a během procesů replikace a rekombinace genů..
Topoizomerázy typu I lze rozdělit na typ 1A a typ 1B. Druhé jsou ty, které se nacházejí u lidí, a jsou zodpovědné za uvolnění nadšroubovicové DNA.
Topoizomeráza 1B (Top1B) je tvořena 765 aminokyselinami rozdělenými do 4 specifických domén. Jedna z těchto domén má vysoce konzervovanou oblast obsahující aktivní místo tyrosinu (Tyr7233). Všechny topoizomerázy představují na svém aktivním místě tyrosin se zásadní rolí v celém katalytickém procesu.
Tyrosin v aktivním místě tvoří kovalentní vazbu s 3'-fosfátovým koncem řetězce DNA, rozřezává jej a udržuje jej připojený k enzymu, zatímco prochází dalším řetězcem DNA štěpením.
Průchod dalšího řetězce DNA děleným řetězcem je dosažen díky konformační transformaci enzymu, který vede k otevření dvojité šroubovice DNA..
Potom se topoizomeráza I vrací do své původní konformace a znovu váže štěpené konce. K tomu dochází procesem obráceným k rozpadu řetězce DNA v katalytickém místě enzymu. Nakonec topoizomeráza uvolní řetězec DNA.
Rychlost ligace DNA je vyšší než rychlost excize, čímž je zajištěna stabilita molekuly a integrita genomu..
Stručně řečeno, topoizomeráza typu I katalyzuje:
Topoizomerázy typu II jsou dimerní enzymy, které štěpí oba řetězce DNA, čímž uvolňují supercoily, které se generují během transkripce a dalších buněčných procesů.
Tyto enzymy potřebují hořčík (Mg++) a také potřebují energii, která pochází z rozpadu trifosfátové vazby ATP, které díky ATPase využívají.
Lidské topoizomerázy II jsou velmi podobné kvasinkám (Saccharomyces cerevisiae), který je tvořen dvěma monomery (podfragmenty A a B). Každý monomer má doménu ATPázy a v dílčím fragmentu aktivní místo tyrosinu 782, na které se může vázat DNA. Dvě vlákna DNA se tedy mohou vázat na topoizomerázu II..
Mechanismus účinku topoizomerázy II je stejný jako mechanismus popsaný pro topoizomerázu I, vzhledem k tomu, že jsou rozštěpeny dva řetězce DNA, a ne pouze jeden..
Na aktivním místě topoizomerázy II je fragment dvojitá spirála DNA, nazývaný „fragment G“. Tento fragment je vyříznut a držen pohromadě k aktivnímu místu kovalentními vazbami..
Poté enzym umožňuje dalšímu fragmentu DNA, nazývanému „T fragment“, projít štěpeným fragmentem „G“, a to díky konformační změně enzymu, která je závislá na hydrolýze ATP..
Topoizomeráza II váže dva konce „fragmentu G“ a nakonec obnoví svůj původní stav uvolněním fragmentu „G“. DNA pak uvolňuje torzní napětí, což umožňuje replikační a transkripční procesy..
Lidský genom má pět topoizomeráz: top1, top3α, top3β (typu I); a top2α, top2β (typu II). Nejrelevantnější lidské topoizomerázy jsou top1 (topoizomeráza typu IB) a 2α (topoizomeráza typu II).
Protože procesy katalyzované topoizomerázami jsou nezbytné pro přežití buněk, jsou tyto enzymy dobrým terčem útoku, který ovlivňuje maligní buňky. Z tohoto důvodu jsou topoizomerázy považovány za důležité při léčbě mnoha lidských onemocnění..
Léky, které interagují s topoizomerázami, jsou v současné době široce studovány jako chemoterapeutické látky proti rakovinovým buňkám (v různých orgánech těla) a patogenním mikroorganismům..
Léky, které inhibují aktivitu topoizomerázy, mohou:
Stabilizace přechodného komplexu, který je tvořen vazbou DNA na tyrosin katalytického místa enzymu, brání vazbě štěpených fragmentů, což může vést k buněčné smrti.
Mezi sloučeniny, které inhibují topoizomerázy, patří následující.
Antibiotika se používají proti rakovině, protože zabraňují růstu nádorových buněk, obvykle interferencí s jejich DNA. Často se jim říká antineoplastická (rakovinová) antibiotika. Actinomycin D například ovlivňuje topoizomerázu II a používá se u Wilmsových nádorů u dětí a rhabdomyosarkomů.
Antracykliny jsou mezi antibiotiky jedním z nejúčinnějších protinádorových léků s nejširším spektrem. Používají se k léčbě rakoviny plic, vaječníků, dělohy, žaludku, močového měchýře, prsu, leukémie a lymfomů. Je známo, že ovlivňuje topoizomerázu II interkalací v DNA.
První antracyklin izolovaný z aktinobakterií (Streptomyces peucetius) byl daunorubicin. Později byl doxorubicin syntetizován v laboratoři a v současné době se také používají epirubicin a idarubicin.
Antrachinony nebo anthracenediony jsou sloučeniny odvozené od antracenu, podobné antracyklinům, které interkalací v DNA ovlivňují aktivitu topoizomerázy II. Používají se pro metastatický karcinom prsu, non-Hodgkinův lymfom (NHL) a leukémii..
Tyto léky byly nalezeny v pigmentech některých druhů hmyzu, rostlin (frangula, senna, rebarbora), lišejníků a hub; stejně jako hoelit, který je přírodním minerálem. V závislosti na vaší dávce mohou být karcinogenní.
Mezi těmito sloučeninami máme mitoxantron a jeho analog, losoxantron. Ty zabraňují šíření maligních nádorových buněk, které se nevratně váží na DNA.
Podofylotoxiny, jako jsou epidophyllotoxiny (VP-16) a teniposid (VM-26), tvoří komplex s topoizomerázou II. Používají se mimo jiné proti rakovině plic, varlat, leukémii, lymfomům, rakovině vaječníků, karcinomu prsu a maligním intrakraniálním nádorům. Jsou izolovány z rostlin Podophyllum notatum Y P. peltatum.
Campotheciny jsou sloučeniny, které inhibují topoizomerázu I, včetně irinotekanu, topotekanu a diflomotekanu..
Tyto sloučeniny se používají proti rakovině tlustého střeva, plic a prsu a získávají se přirozeně z kůry a listů stromových druhů. Camptotheca acuminata čínských corniceas a Tibetu.
Ke strukturním změnám topoizomeráz I a II může dojít také zcela přirozeně. K tomu může dojít během některých událostí, které ovlivňují váš katalytický proces.
Mezi tyto změny patří tvorba pyrimidinových dimerů, neshody bází dusíku a další události způsobené oxidačním stresem..



Zatím žádné komentáře