
The Golgiho aparát, Také známý jako Golgiho komplex, je to membránová buněčná organela tvořená sadou plochých vezikul skládaných dohromady; Tyto vaky mají uvnitř tekutinu. Nalezeno v široké škále eukaryot, včetně zvířat, rostlin a hub.
Tato organela je zodpovědná za zpracování, balení, klasifikaci, distribuci a modifikaci proteinů. Kromě toho má také roli v syntéze lipidů a sacharidů. Na druhé straně v rostlinách dochází k syntéze složek buněčné stěny v Golgiho aparátu..
Golgiho aparát byl objeven v roce 1888, zatímco byly studovány nervové buňky; jeho objevitel, Camillo Golgi, získal Nobelovu cenu. Struktura je detekovatelná barvením chromátem stříbrným.
Zpočátku byla existence orgánu pro tehdejší vědce pochybná a přisuzovali Golgiho pozorování jednoduchým artefaktům produktům použitých technik.
Rejstřík článků
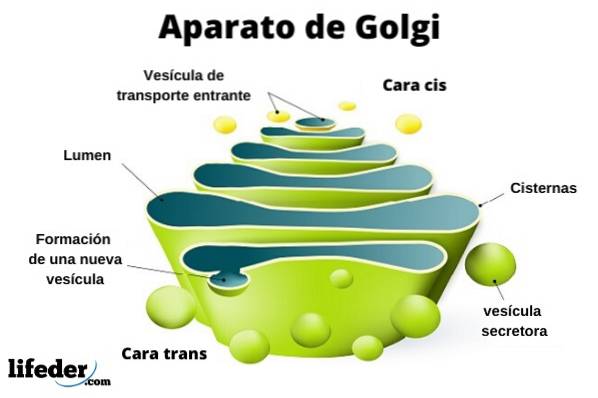
Golgiho aparát je eukaryotická organela membránové povahy. To se podobá taškám v hromadách, i když organizace se může lišit v závislosti na typu buňky a organismu. Je zodpovědný za modifikaci proteinů po translaci.
Například může být přidán nějaký sacharid za vzniku glykoproteinu. Tento produkt je zabalen a distribuován do buněčného prostoru, kde je to požadováno, jako je membrána, lysozomy nebo vakuoly; lze jej také odeslat mimo buňku. Podílí se také na syntéze biomolekul.
Cytoskelet (konkrétně aktin) určuje jeho umístění a komplex se obecně nachází v oblasti vnitřku buňky v blízkosti jádra a centrosomu.
Golgiho komplex je tvořen sadou plochých, fenestrovaných diskovitých vaků, zvaných golgické cisterny, různé tloušťky.
Tyto pytle jsou naskládané do skupin po čtyřech nebo šesti cisternách. V savčí buňce najdete mezi 40 a 100 vzájemně propojenými bateriemi.
Golgiho komplex představuje zajímavou vlastnost: existuje polarita z hlediska struktury i funkce.
Můžete rozlišovat mezi cis tváří a trans tváří. První souvisí se vstupem proteinů a nachází se v blízkosti endoplazmatického retikula. Druhým je výstupní plocha nebo sekrece produktu; Jsou tvořeny jednou nebo dvěma nádržemi, které mají trubkovitý tvar.
Spolu s touto strukturou jsou vezikuly, které tvoří dopravní systém. Hromady pytlů jsou navzájem spojeny strukturou připomínající luk nebo datum.
U savců je komplex Golgi během procesů dělení buněk fragmentován na několik vezikul. Vezikuly přecházejí do dceřiných buněk a znovu mají tradiční podobu komplexu.
Organizace komplexu není u všech skupin organismů běžná. V některých typech buněk není komplex strukturován jako sady cisteren naskládaných do skupin; naopak, jsou umístěny jednotlivě. Příkladem této organizace je houba Saccharomyces cerevisiae.
V některých jednobuněčných organismech, jako je toxoplazma nebo trypanosoma, byla hlášena přítomnost pouze jedné membránové hromádky.
Všechny tyto výjimky naznačují, že stohování struktur není nezbytné pro splnění jejich funkce, ačkoli blízkost mezi vaky činí přepravní proces mnohem efektivnějším..
Podobně některým bazálním eukaryotům tyto cisterny chybí; například houby. Tento důkaz podporuje teorii, že se přístroj objevil v pozdější linii než první eukaryoti..
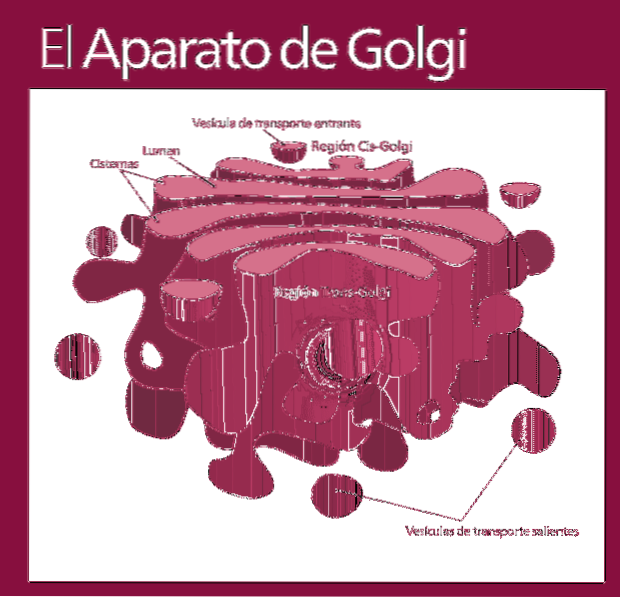
Komplex Golgi je funkčně rozdělen do následujících oddílů: síť cis, naskládané vaky - které jsou zase rozděleny na střední a trans subkompartment - a trans.
Molekuly, které budou modifikovány, vstupují do Golgiho komplexu ve stejném pořadí (cis síť, následovaná dílčími částmi, které budou nakonec vyloučeny v trans síti).
Většina reakcí probíhá v nejaktivnějších zónách: trans a střední subpartpartments.
Golgiho komplex má jako hlavní funkci posttranslační modifikaci proteinů díky enzymům, které mají uvnitř.
Tyto modifikace zahrnují procesy glykosylace (přidání sacharidů), fosforylace (přidání fosfátové skupiny), sulfatace (přidání fosfátové skupiny) a proteolýzy (degradace proteinů).
Golgiho komplex se dále podílí na syntéze specifických biomolekul. Každá z jejích funkcí je podrobně popsána níže:
V Golgiho aparátu dochází k modifikaci proteinu na glykoprotein. Typické kyselé pH vnitřku organely je kritické pro to, aby tento proces probíhal normálně..
Mezi Golgiho aparátem a endoplazmatickým retikulem a lysozomy dochází k neustálé výměně materiálů. V endoplazmatickém retikulu procházejí proteiny také úpravami; mezi ně patří přidání oligosacharidu.
Když tyto molekuly (N-oligosacharidy) vstoupí do Golgiho komplexu, dostanou řadu dalších modifikací. Pokud má být osud této molekuly přenesen mimo buňku nebo má být přijat v plazmatické membráně, nastanou speciální úpravy.
Tyto modifikace zahrnují následující kroky: odstranění tří zbytků manózy, přidání N-acetylglukosaminu, odstranění dvou manóz a přidání fukózy, dvou dalších N-acetylglukosaminu, tří galaktózy a tří zbytků kyseliny sialové..
Naproti tomu proteiny, které jsou určeny pro lysozomy, jsou modifikovány následujícím způsobem: v počátečním kroku nedochází k odstranění manóz; místo toho dochází k fosforylaci těchto zbytků. K tomuto kroku dochází v cis oblasti komplexu.
Dále se odstraní N-acetylglukosaminové skupiny, přičemž se manózy ponechají s fosfátem přidaným k oligosacharidu. Tyto fosfáty naznačují, že protein musí být specificky zaměřen na lysozomy..
Receptory, které jsou zodpovědné za rozpoznávání fosfátů, které indikují jejich intracelulární osud, se nacházejí v trans síti.
V komplexu Golgi dochází k syntéze glykolipidů a sfingomyelinu za použití ceramidu (dříve syntetizovaného v endoplazmatickém retikulu) jako molekuly původu. Tento proces je v rozporu s procesem u ostatních fosfolipidů, které tvoří plazmatickou membránu a které jsou odvozeny od glycerolu..
Sfingomyelin je třída sfingolipidů. Je hojnou složkou savčích membrán, zejména nervových buněk, kde jsou součástí myelinového obalu.
Po jejich syntéze jsou transportovány do svého konečného umístění: plazmatické membrány. Jejich polární hlavy jsou umístěny směrem ven z povrchu buňky; tyto prvky mají specifickou roli v procesech rozpoznávání buněk.
V rostlinných buňkách přispívá Golgiho aparát k syntéze polysacharidů, které tvoří buněčnou stěnu, konkrétně hemicelulózy a pektinů. Vezikulárním transportem jsou tyto polymery přenášeny mimo buňku.
U zeleniny je tento krok zásadní a přibližně 80% aktivity retikula je přiřazeno syntéze polysacharidů. Ve skutečnosti byly stovky těchto organel popsány v rostlinných buňkách..
Různé biomolekuly - bílkoviny, sacharidy a lipidy - jsou přenášeny do svých buněčných destinací komplexem Golgi. Proteiny mají jakýsi „kód“, který je zodpovědný za informování cíle, ke kterému patří.
Jsou transportovány ve váčcích, které opouštějí trans síť a cestují do specifického buněčného kompartmentu.
Proteiny mohou být přeneseny na membránu specifickou konstitutivní cestou. Proto dochází k kontinuální inkorporaci proteinů a lipidů do plazmatické membrány. Tím jsou zachovány proteiny, jejichž konečným cílem je komplex Golgi.
Kromě konstitutivní dráhy jsou další proteiny určeny pro vnějšek buňky a vyskytují se prostřednictvím signálů z prostředí, ať už jde o hormony, enzymy nebo neurotransmitery..
Například v pankreatických buňkách jsou trávicí enzymy zabaleny do vezikul, které se vylučují pouze při detekci přítomnosti potravy..
Nedávný výzkum uvádí existenci alternativních drah membránových proteinů, které neprocházejí Golgiho aparátem. Tyto cesty však bypass O „nekonvenčních“ se v literatuře diskutuje.
Existuje pět modelů vysvětlujících obchodování s proteiny v přístroji. První zahrnuje přenos materiálu mezi stabilními oddíly, každý z nich má nezbytné enzymy pro splnění konkrétních funkcí. Druhý model zahrnuje postupné zrání cisteren.
Třetí také navrhuje zrání sáčků, ale se začleněním nové komponenty: trubkové dopravy. Podle modelu jsou tubuly důležité v provozu v obou směrech.
Čtvrtý model navrhuje, aby komplex fungoval jako celek. Pátý a poslední model je nejnovější a tvrdí, že komplex je rozdělen do různých oddílů.
V určitých typech buněk má komplex Golgiho specifické funkce. Buňky pankreatu mají speciální struktury pro sekreci inzulínu.
Různé typy krve u lidí jsou příkladem diferenciálních glykosylačních vzorců. Tento jev je vysvětlen přítomností různých alel, které kódují glukotransferázu.



Zatím žádné komentáře