
The Genetický drift nebo gen je stochastický evoluční mechanismus, který způsobuje fluktuace nebo čistě náhodné variace alelových frekvencí populace.
Přirozený výběr a drift genů Charlese Darwina jsou dva nejdůležitější procesy, které se podílejí na evoluční změně v populacích. Na rozdíl od přirozeného výběru, který je považován za deterministický a nenáhodný proces, je drift genů proces, který se prokazuje jako náhodné fluktuace frekvencí alel v populaci nebo haplotypů..
Genový drift vede k neadaptivní evoluci. Ve skutečnosti je přirozený výběr - a nikoli genetický drift - jediným mechanismem používaným k vysvětlení všech adaptací organismů na různých úrovních (anatomických, fyziologických nebo etologických)..
To neznamená, že drift genu není důležitý. Jeden z nejvýraznějších důsledků tohoto jevu je pozorován na molekulární úrovni, mezi rozdíly v DNA a proteinových sekvencích.
Rejstřík článků
Teorie driftu genů byla vyvinuta na počátku 30. let významným biologem a genetikem jménem Sewal Wright..
Stejně tak byly příspěvky Motoo Kimury v této oblasti výjimečné. Tento výzkumník vedl neutrální teorii molekulární evoluce, kde odhaluje, že účinky driftu genů mají důležitý příspěvek k evoluci na úrovni sekvencí DNA..
Tito autoři navrhli matematické modely, aby pochopili, jak funguje drift genů v biologických populacích..
Příčiny driftu genů jsou stochastické jevy - tedy náhodné. Ve světle populační genetiky je evoluce definována jako časová variace ve frekvencích alel populace. Drift vede ke změně těchto frekvencí náhodnými událostmi zvanými „chyby vzorkování“.
Genový drift je považován za chybu vzorkování. Geny, které jsou zahrnuty v každé generaci, jsou vzorkem genů nesených předchozí generací.
Jakýkoli vzorek podléhá chybě při odběru vzorků. To znamená, že podíl různých položek, které najdeme ve vzorku, se může změnit čistou náhodou..
Představme si, že máme tašku s 50 bílými žetony a 50 černými žetony. Vezmeme-li deset z nich, možná čistou náhodou získáme 4 bílé a 6 černých; nebo 7 bílých a 3 černé. Existuje rozpor mezi teoreticky očekávanými hodnotami (5 a 5 každé barvy) a hodnotami získanými experimentálně..
Účinky driftu genů se projevují jako náhodné změny ve frekvencích alel populace. Jak jsme zmínili, k tomu dochází, když neexistuje žádný vztah mezi charakteristikou, která se mění, a zdatnost. Postupem času budou alely nakonec z populace fixovány nebo ztraceny.
V evoluční biologii, termín zdatnost Je široce používán a odkazuje na schopnost organismu reprodukovat se a přežít. Parametr se pohybuje mezi 0 a 1.
Charakteristika, která se mění podle driftu, tedy nesouvisí s reprodukcí a přežitím jedince..
Ztráta alel vede k druhému účinku driftu genů: ke ztrátě heterozygotnosti v populaci. Variace na určitém místě se sníží a nakonec bude ztracena.
Pravděpodobnost, že bude alela fixována v populaci, se rovná její frekvenci v době, kdy je studována. Četnost připojení alternativní alely bude 1 - p. Kde p se rovná frekvenci alely.
Na tuto frekvenci nemá vliv předchozí historie změn ve frekvencích alel, takže nelze ani předpovídat na základě minulosti..
Pokud alela naopak vznikla mutací, je její pravděpodobnost fixace p = 1/2N. Kde N je počet obyvatel. To je důvod, proč se nové alely, které se objevují mutací, snadněji fixují u malých populací..
Čtenář musí zdůvodnit, jak je hodnota p když je jmenovatel menší. Logicky by se pravděpodobnost zvýšila.
Účinky driftu genů tedy probíhají rychleji u malých populací. V diploidní populaci (dvě sady chromozomů, jako my lidé) dochází k připojení nových alel v průměru každé 4N generace. Čas roste úměrně s růstem N z populace.
The N který se objevuje v předchozích rovnicích, neodkazuje na hodnotu shodnou s počtem jednotlivců, kteří tvoří populaci. To znamená, že to není ekvivalentní sčítání organismů.
V populační genetice je parametr „efektivní počet obyvatel“ (Ne), což je obvykle méně než u všech jednotlivců.
Například u některých populací se sociální strukturou, v níž dominuje pouze několik mužů, je efektivní počet obyvatel velmi nízký, protože geny těchto dominantních mužů nepřiměřeně přispívají - pokud je porovnáme se zbytkem mužů..
Z tohoto důvodu bude rychlost, s jakou genový drift působí (a rychlost, se kterou se ztrácí heterozygosita) větší, než jsme očekávali, pokud provedeme sčítání, protože populace je menší, než se zdá..
Pokud v hypotetické populaci počítáme 20 000 jedinců, ale pouze 2 000 se množí, efektivní počet populace klesá. A tento jev, ve kterém se v populaci nevyskytují všechny organismy, je široce rozšířen v přírodních populacích..
Jak jsme zmínili (a matematicky se prokázali), drift se vyskytuje u malých populací. Kde jsou alely, které nejsou tak časté, pravděpodobněji ztraceny.
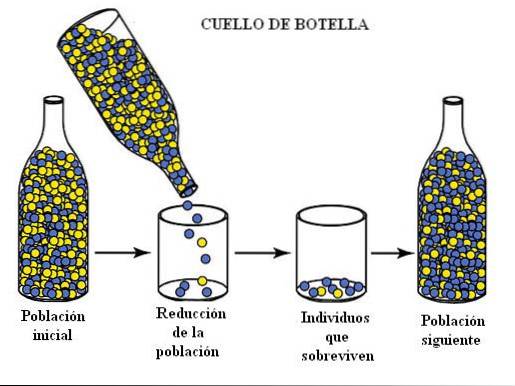
Tento jev je běžný poté, co populace zažívá událost zvanou „úzké místo“. K tomu dochází, když je značný počet členů populace eliminován nějakým typem nepředvídané nebo katastrofické události (například bouře nebo lavina).
Okamžitým účinkem by mohlo být snížení genetické rozmanitosti populace, zmenšení velikosti genofondu nebo genofondu.
Specifickým případem úzkých míst je efekt zakladatele, kdy se malý počet jedinců odděluje od původní populace a vyvíjí se izolovaně. Na příkladech, které uvedeme později, uvidíme, jaké jsou důsledky tohoto jevu.
Neutrální teorii molekulární evoluce navrhl Motoo Kimura. Před myšlenkami tohoto výzkumníka již Lewontin & Hubby zjistili, že vysoký podíl variací na úrovni enzymu nemůže aktivně udržovat všechny tyto polymorfismy (variace).
Kimura dospěl k závěru, že tyto změny aminokyselin lze vysvětlit driftem genů a mutacemi. Došel k závěru, že na úrovni DNA a bílkovin hrají mechanismy driftu genů zásadní roli..
Termín neutrální označuje skutečnost, že většina základních substitucí, které se podaří zafixovat (dosáhnout frekvence 1), je neutrální vzhledem k zdatnost. Proto tyto variace, ke kterým dochází driftem, nemají žádný adaptivní význam..
Existují mutace, které nemají vliv na fenotyp jedince. Veškeré informace o vývoji a vývoji nového organismu jsou šifrovány v DNA. Tento kód je v procesu překladu dešifrován ribozomy..
Genetický kód se čte v „tripletech“ (sada tří písmen) a každé tři písmena kódují aminokyselinu. Genetický kód je však zdegenerovaný, což naznačuje, že existuje více než jeden kodon, který kóduje stejnou aminokyselinu. Například kodony CCU, CCC, CCA a CCG kódují aminokyselinu prolin..
Pokud se tedy sekvence CCU změní na CCG, translačním produktem bude prolin a v proteinové sekvenci nedojde k žádným modifikacím..
Podobně se mutace může změnit na aminokyselinu, jejíž chemické vlastnosti se příliš neliší. Například pokud se alanin změní na valin možná účinek na funkčnost proteinu je nepostřehnutelný.
Toto neplatí ve všech případech, pokud ke změně dojde v části proteinu, která je nezbytná pro jeho funkčnost - jako je aktivní místo enzymů - účinek na zdatnost může to být velmi významné.
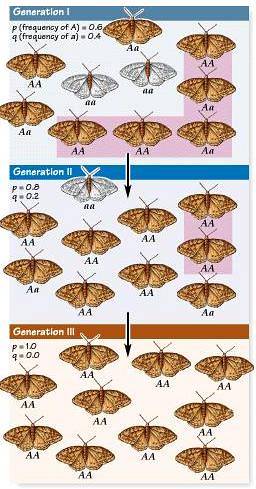
Představte si louku, kde koexistují hlemýždi a krávy. V populaci hlemýžďů můžeme rozlišit dvě zbarvení: černou skořápku a žlutou skořápku. Rozhodujícím faktorem úmrtnosti hlemýžďů jsou stopy krav.
Mějte však na paměti, že pokud se na šneka vstoupí, nezávisí to na barvě skořápky, protože jde o náhodnou událost. V tomto hypotetickém příkladu začíná populace hlemýžďů se stejným podílem barev (50 černých hlemýžďů a 50 žlutých hlemýžďů). V případě krav eliminujte 6 černých a pouze 2 žluté, podíl barev by se změnil.
Stejným způsobem může dojít k tomu, že v následujícím případě zemřou žluté ve větším poměru, protože neexistuje žádný vztah mezi barvou a pravděpodobností rozdrcení (neexistuje však žádný typ „kompenzačního“ efektu).
Během tohoto náhodného procesu budou proporce černé a žluté skořápky v průběhu času kolísat. Nakonec jedna ze skořápek dosáhne jednoho ze dvou limitů: 0 nebo 1.
Když je dosažená frekvence 1 - předpokládejme alelu žluté skořápky - všichni hlemýždi budou mít tuto barvu. A jak můžeme uhodnout, alela černé skořápky bude ztracena.
Jediným způsobem, jak tuto alelu znovu získat, je populace, která vstupuje migrací nebo mutací.
Fenomén driftu genů lze pozorovat v přírodních populacích a nejextrémnějším příkladem jsou gepardi. Tyto rychlé a stylové kočkovité šelmy patří k tomuto druhu Acinonyx jubatus.
Asi před 10 000 lety zažili gepardi - a další velké populace savců - událost extrémního vyhynutí. Tato událost způsobila „úzké místo“ v populaci gepardů, kde přežilo jen několik jedinců..
Ti, kdo přežili pleistocénní katastrofický jev, dali vzniknout všem současným gepardům. Účinky driftu spolu s příbuzenským plemenitbou způsobily téměř úplnou homogenizaci populace.
Ve skutečnosti je imunitní systém těchto zvířat u všech jedinců prakticky totožný. Pokud některý z členů z nějakého důvodu potřeboval darování orgánu, mohl by tak učinit kterýkoli z jeho kolegů, aniž by to vedlo k jakékoli pravděpodobnosti odmítnutí..
Dary jsou postupy, které jsou pečlivě prováděny a je nutné potlačit imunitní systém příjemce tak, aby nenapadl „vnějšího agenta“, i když pochází od velmi blízkého příbuzného - říkejte tomu bratři nebo děti..
Úzká místa a efekt zakladatele se vyskytují také u současné lidské populace a mají velmi relevantní důsledky v lékařské oblasti.
Amiši jsou náboženská skupina. Vyznačují se jednoduchým životním stylem bez technologií a dalšího současného pohodlí - navíc s extrémně vysokou frekvencí nemocí a genetických patologií.
Asi 200 kolonizátorů dorazilo do Pensylvánie (USA) z Evropy a začalo se rozmnožovat mezi stejnými členy.
Předpokládá se, že mezi kolonizátory byli nositelé autozomálně recesivních genetických onemocnění, včetně Ellis-van Creveldova syndromu. Tento syndrom je charakterizován rysy nanismu a polydaktylie (vysoký počet prstů, více než pět číslic).
Nemoc byla v počáteční populaci s frekvencí 0,001 a významně vzrostla na 0,07.



Zatím žádné komentáře