The anaerobní glykolýza nebo anaerobní je katabolická cesta používaná mnoha typy buněk k rozpadu glukózy v nepřítomnosti kyslíku. To znamená, že glukóza není úplně oxidována na oxid uhličitý a vodu, jako je tomu v případě aerobní glykolýzy, ale vznikají fermentační produkty..
Nazývá se anaerobní glykolýza, protože probíhá bez přítomnosti kyslíku, který v ostatních případech funguje jako konečný akceptor elektronů v mitochondriálním transportním řetězci, kde se při zpracování glykolytických produktů produkuje velké množství energie..
V závislosti na organismu bude mít stav anaerobiózy nebo nepřítomnosti kyslíku za následek produkci kyseliny mléčné (například svalové buňky) nebo ethanolu (kvasinky) z pyruvátu generovaného katabolismem glukózy.
Výsledkem je, že výtěžek energie drasticky klesá, protože na každý zpracovávaný mol glukózy se produkují pouze dva moly ATP, ve srovnání s 8 moly, které lze získat během aerobní glykolýzy (pouze v glykolytické fázi)..
Rozdíl v počtu molekul ATP souvisí s reoxidací NADH, která negeneruje další ATP, na rozdíl od toho, co se děje v aerobní glykolýze, kde se pro každý NADH získají 3 molekuly ATP.
Rejstřík článků
Anaerobní glykolýza se vůbec neliší od aerobní glykolýzy, protože termín „anaerobní“ odkazuje spíše na to, co se stane po glykolytické cestě, tj. Na osud reakčních produktů a meziproduktů..
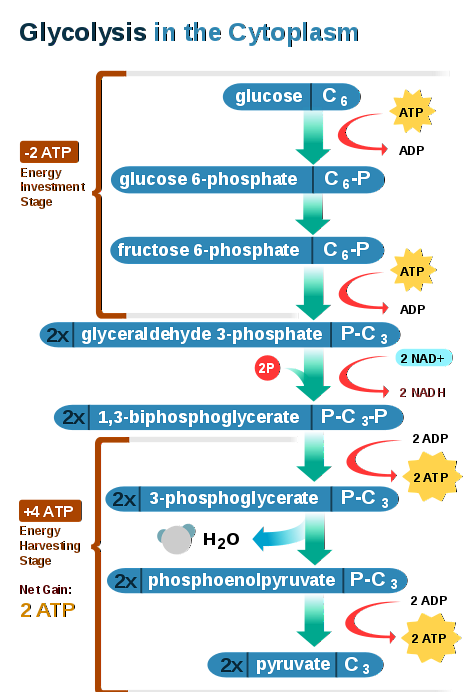
Na anaerobních glykolýzních reakcích se tedy podílí deset různých enzymů, jmenovitě:
1-Hexokináza (HK): používá jednu molekulu ATP pro každou molekulu glukózy. Produkuje 6-fosfát glukózy (G6P) a ADP. Reakce je nevratná a vyžaduje ionty hořčíku.
2-fosfoglukóza izomeráza (PGI): izomerizuje G6P na fruktóza 6-fosfát (F6P).
3-Fosfofruktokináza (PFK): fosforyluje F6P na 1,6-bisfosfát fruktózy (F1,6-BP) pomocí jedné molekuly ATP pro každý F6P, tato reakce je také nevratná.
4-Aldoláza: štěpí molekulu F1,6-BP a produkuje glyceraldehyd-3-fosfát (GAP) a dihydroxyacetonfosfát (DHAP).
5-triosa fosfát izomeráza (TIM): podílí se na interkonverzi DHAP a GAP.
6-glyceraldehyd-3-fosfátdehydrogenáza (GAPDH): používá dvě molekuly NAD+ a 2 molekuly anorganického fosfátu (Pi) k fosforylaci GAP, výtěžek 1,3-bisfosfoglycerát (1,3-BPG) a 2 NADH.
7-fosfoglycerátkináza (PGK): produkuje dvě molekuly ATP fosforylací na úrovni substrátu dvou molekul ADP. Jako dárce fosfátové skupiny používá každou molekulu 1,3-BPG. Produkuje 2 molekuly 3-fosfoglycerátu (3PG).
8-fosfoglycerát mutáza (PGM): přeskupí molekulu 3PG a vytvoří meziprodukt s vyšší energií, 2PG.
9-enoláza: z 2PG produkuje fosfoenolpyruvát (PEP) dehydratací prvního.
10-Pyruvátkináza (PYK): fosforenolpyruvát je používán tímto enzymem k tvorbě pyruvátu. Reakce zahrnuje přenos fosfátové skupiny v poloze 2 z fosfoenolpyruvátu na molekulu ADP. Pro každou glukózu se vytvoří 2 pyruváty a 2 ATP.
Fermentace je termín používaný k označení, že glukóza nebo jiné živiny jsou degradovány v nepřítomnosti kyslíku za účelem získání energie.
V nepřítomnosti kyslíku nemá elektronový transportní řetězec konečný akceptor, a proto nedochází k oxidativní fosforylaci, která vede k velkému množství energie ve formě ATP. NADH není reoxidován mitochondriální cestou, ale alternativními cestami, které neprodukují ATP.
Nestačí NAD+ glykolytická cesta je zastavena, protože přenos fosfátu na GAP vyžaduje současnou redukci tohoto kofaktoru.
Některé buňky mají alternativní mechanismy pro zvládnutí období anaerobiózy a obecně tyto mechanismy zahrnují určitý typ fermentace. Na druhou stranu jiné buňky téměř výhradně závisí na fermentačních procesech..
Produkty fermentačních cest mnoha organismů jsou pro člověka ekonomicky relevantní; Příkladem je produkce ethanolu některými anaerobními kvasinkami a tvorba kyseliny mléčné lakto-bakteriemi používanými k výrobě jogurtu..
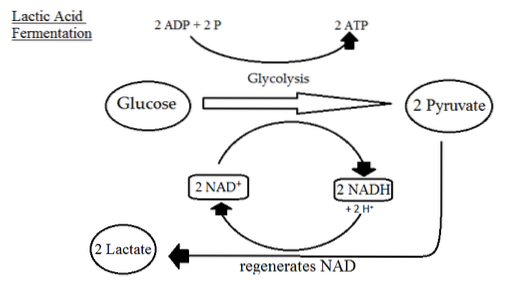
Mnoho typů buněk v nepřítomnosti kyslíku produkuje kyselinu mléčnou díky reakci katalyzované komplexem laktátdehydrogenázy, který využívá uhlíky pyruvátu a NADH produkovaný v GAPDH reakci..
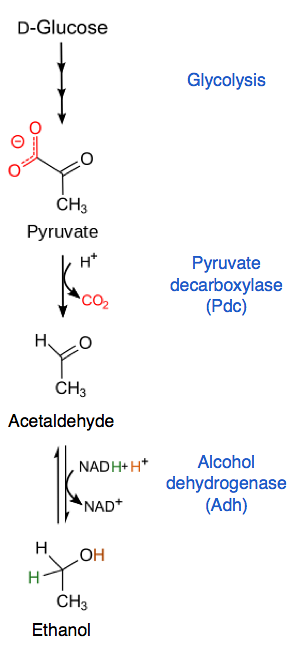
Pyruvát se převádí na acetaldehyd a CO2 pyruvát dekarboxylázou. Acetaldehyd je poté používán alkohol dehydrogenázou, která ho redukuje, produkuje ethanol a regeneruje molekulu NAD.+ pro každou molekulu pyruvátu, která vstupuje tímto způsobem.
Anaerobní glykolýza má jako hlavní charakteristiku skutečnost, že konečné produkty neodpovídají COdva a voda, jako v případě aerobní glykolýzy. Místo toho se generují typické produkty fermentačních reakcí.
Někteří autoři popsali proces „aerobní fermentace“ nebo aerobní glykolýzy glukózy pro určité organismy, mezi nimiž vynikají někteří paraziti z čeledi Trypanosomatidae a mnoho rakovinných nádorových buněk..
U těchto organismů se ukázalo, že i v přítomnosti kyslíku produkty glykolytické dráhy odpovídají produktům fermentačních cest, a proto se má za to, že dochází k „částečné“ oxidaci glukózy, protože ne veškerá energie je možné uhlíky.
Ačkoli „aerobní fermentace“ glukózy neznamená úplnou absenci respirační aktivity, protože nejde o proces všeho nebo nic. Literatura však naznačuje vylučování produktů, jako jsou pyruvát, laktát, sukcinát, malát a další organické kyseliny..
Mnoho rakovinných buněk vykazuje zvýšení absorpce glukózy a glykolytického toku.
Nádory u pacientů s rakovinou rychle rostou, takže cévy jsou hypoxické. Energetický doplněk těchto buněk tedy závisí hlavně na anaerobní glykolýze..
Tomuto jevu však napomáhá transkripční faktor indukovaný hypoxií (HIF), který prostřednictvím komplexních mechanismů zvyšuje expresi glykolytických enzymů a transportérů glukózy v membráně..


Zatím žádné komentáře