The askospory jsou výtrusy produktem sexuální reprodukce, karyogamie a meiotického dělení v asci hub třídy Ascomycota. Ty, když vyklíčí, mohou způsobit vznik nové nezávislé houby.
Ascomycota nebo Ascomycetes jsou třídou hub, která zahrnuje asi 30% všech známých druhů hub. Vyskytují se nejhojněji v suchozemském a vodním prostředí. Pro mořská stanoviště je typických pouze několik druhů.
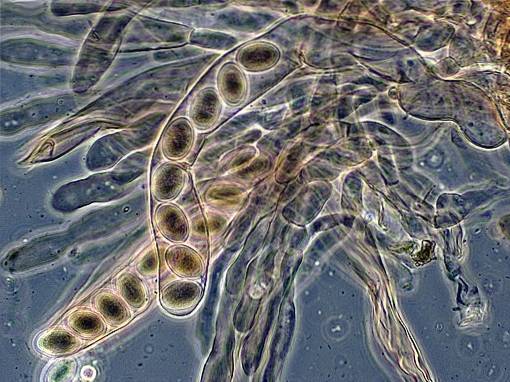
Charakteristickým rysem ascomycetes je vytvoření struktury produkující endospory. Tato struktura představuje speciální typ sporangia a nazývá se „znechucení“. Proto jsou všechny houby, které způsobují znechucení, třídou Ascomycetes..
Asci mají obecně vakovitý tvar a představují místo, kde se tvoří askospory. Specializovanější ascomycety, jako jsou ty, které se vyskytují u lišejníků, mají makroskopické asci a plodnice zvanou askokarpus..
Tvar asci a ascospores používají taxonomové k rozlišení různých druhů třídy Ascomycota. Například v ascomycetách jsou kvasinky, jednobuněčné houby, které netvoří plodnice..
Část zemědělsko-potravinářského průmyslu se věnuje konzervování předmětů a potravin před kontaminací askospory, protože když vyklíčí a vzniknou dospělí jedinci, zhoršují a rozkládají potraviny.
Rejstřík článků
Askospory jsou jako jakési „semeno“ hub Ascomycota, obdobné jako u rostlin, protože mohou zůstat neaktivní (v klidu), ale žít po dlouhou dobu..
Tyto struktury jsou velmi odolné, mohou vést k novým úplným jedincům a mohou zůstat naživu ještě dlouho po vyklíčení, protože se živí endogenními substráty..
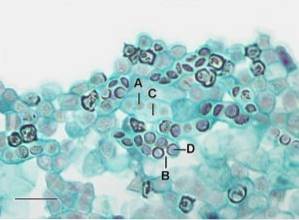
Askospory však mají jedinečné vlastnosti, které je odlišují od semen rostlin, například hlavním stimulem pro klíčení askospor jsou chemické látky produkované rozkladem substrátů..
U rostlin je naopak stimulací pro klíčení v některých případech voda a světlo. Askospory mají polovinu chromozomálního náboje normální buňky, to znamená, že jsou haploidní; zatím jsou semena rostlin většinou polyploidní.
Askospory jsou obecně mikroskopické struktury, které jsou zřídka mírně viditelné s lupami s malým zvětšením. Na druhou stranu jsou semena zeleniny makroskopická a u mikroskopických semen lze pojmenovat několik výjimek..
Když se díváme pod mikroskop a podrobně popisujeme typickou askosporu, pozorujeme, že mají eliptický tvar, že mají své protoplasty uzavřené tříúrovňovou nebo vrstvenou chitinovou buněčnou stěnou a že mají zárodečný pór na každém konci buňky..
Askospory jsou konečným produktem procesu sexuální reprodukce ascomycet. Tvorba mycelií v těchto organismech začíná klíčením askospor a hned poté se začnou tvořit konidiofory..
Houba začíná růstovou fázi, kdy se produkuje velké množství konidií, které přispívají k šíření houby v substrátu. V tomto myceliu začíná tvorba znechucení.
Před tím dochází k gametogenezi, při které se tvoří anteridie (mužské) a ascogonia (ženské). Jádra antheridia jsou přenesena do ascogonia a protoplasty obou buněk fúzují v procesu zvaném plazmogamie..
Ve stejném cytosolu se samčí jádra páří s ženskými jádry, ale bez fúze. Potom „hyfální“ vlákna začnou růst mimo ascogonium a askogenní hyfy se prodlužují.
V askogenních hyfách se jádra vyvíjejí a množí simultánními mitotickými děleními ve všech hyfách askogonia. Znechucení se tvoří na konci jedné z aschogenních dikaryotických hyf, které vznikly během tohoto kroku..
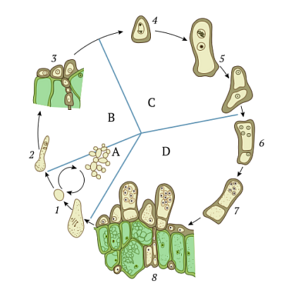
Jedna z buněk dikaryotických hyf roste a vytváří háček zvaný „uncinculus“. V této buňce ve tvaru háku se obě jádra dělí tak, že jejich mitotická vřetena jsou uspořádána v paralelní a vertikální orientaci..
Dvě z dceřiných jader jsou v horní oblasti háku, jedno je blízko konce a druhé poblíž bazální přepážky háku. Existují dvě přepážky, které rozdělují hák na tři buňky.
Buňka uprostřed tří je ta, která způsobí znechucení. Uvnitř této buňky dochází k procesu karyogamie, kdy se obě jádra spojí a vytvoří diploidní jádro známé jako zygota..
Toto diploidní jádro je jediným diploidem v životním cyklu hub Ascomycota. Po karyogamii začíná znechucení dozrávat a prodlužovat se (prodlužovat se).
V mladých buňkách asco podléhají diploidní jádra v nich meióze a později mitóze. Z původní buňky pochází 8 nových haploidních buněk. Těchto osm buněk, jak se vyvinou, se promění v askospory.
Každé jádro, které vzniklo z meiotické a později mitotické reprodukce, bude uloženo společně s částí cytosolu buňky, kde došlo k rozdělení, v buněčné stěně chitinu, která je syntetizována uvnitř buňky..
Téměř ve všech ascomycetách je znechucení velmi dobře strukturovanou tuhou strukturou. Jak askospory dozrávají, znechucení se rozpadá a uvolňuje askospory do prostředí..
Obecně se askospory šíří na krátké vzdálenosti, kolem několika centimetrů, u některých druhů se však šíří až na několik metrů, vše záleží na prostředí, kde jsou vyhnáni..
Nejhojnějším druhem Ascomycoty v přírodě a zemědělství jsou kvasinky, které se nacházejí na povrchu půdy, vody, ovoce a velkého množství potravy..
Tyto organismy mají schopnost metabolizovat cukry, přičemž při tomto procesu produkují alkohol a oxid uhličitý..
Plodnice se v kvasinkách nevyskytují, protože se jedná o jednobuněčné organismy, které se množí nejčastěji binárním štěpením nebo pučením. Pokud jsou však podmínky v médiu nepříznivé, dvě kompatibilní buňky fúzují a tvoří zygotu..
Zygota se vyvíjí přímo uvnitř buňky, tato buňka se diferencuje na odpor a uvnitř se dělí 4 nebo 8 jader v závislosti na druhu kvasinek. Tato jádra se vyvíjejí a jsou pokryta chitinem a transformují se do askospor.
Všechny houby, které tvoří symbiotické sdružení, které představuje lišejníky jsou z čeledi Ascomycota, proto si askospory vyvíjí prostřednictvím své sexuální reprodukce.
Obecně platí, že při podrobném pozorování lišejníku, který již dosáhl svého stádia dospělosti, lze vidět malé miskovité struktury. Tyto struktury jsou plodnice houby, známé jako „apothecia“. Uvnitř apotécie je místo, kde se generují askospory.



Zatím žádné komentáře