
The buněčné dělení je to proces, který umožňuje všem živým organismům růst a množit se. U prokaryot a eukaryot je výsledkem buněčného dělení dceřiné buňky, které mají stejnou genetickou informaci jako původní buňka. Stává se to proto, že před rozdělením jsou informace obsažené v DNA duplikovány.
U prokaryot dochází k dělení binárním štěpením. Genom většiny prokaryot je kruhová molekula DNA. Ačkoli tyto organismy nemají jádro, DNA je v kompaktní formě, která se nazývá nukleoid, která se liší od cytoplazmy, která ji obklopuje..
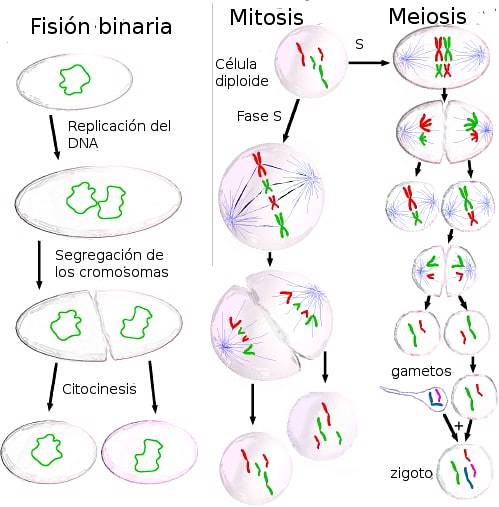
U eukaryot se dělení děje prostřednictvím mitózy a meiózy. Eukaryotický genom se skládá z velkého množství DNA uspořádané v jádře. Tato organizace je založena na balení DNA s proteiny, které tvoří chromozomy, které obsahují stovky nebo tisíce genů..
Velmi různorodé eukaryoty, jednobuněčné i metazoánské, mají životní cykly, které se střídají mezi mitózou a meiózou. Jedná se o cykly s: a) gametickou meiózou (zvířata, některé houby a řasy), b) zygotickou meiózou (některé houby a prvoky); ac) střídání gametické a zygotické meiózy (rostliny).
Rejstřík článků
Buněčné dělení může být binární štěpení, mitóza nebo meióza. Každý z procesů zapojených do těchto typů buněčného dělení je popsán níže..
Binární štěpení se skládá z rozdělení buňky, které vede ke vzniku dvou dceřiných buněk, z nichž každá má identickou kopii DNA původní buňky.
Před dělením prokaryotických buněk dochází k replikaci DNA, která začíná na konkrétním místě dvouvláknové DNA, které se nazývá počátek replikace. Replikační enzymy se pohybují v obou směrech od původu a vytvářejí jednu kopii každého z řetězců dvouvláknové DNA..
Po replikaci DNA se buňka prodlužuje a DNA se v buňce oddělí. Okamžitě začne uprostřed buňky růst nová plazmatická membrána, která vytvoří přepážku..
Tento proces usnadňuje protein FtsZ, který je evolučně vysoce konzervovaný u prokaryot, včetně Archea. Nakonec se buňka rozdělí.
Fáze, kterými eukaryotická buňka prochází mezi dvěma po sobě následujícími buněčnými děleními, se nazývají buněčný cyklus. Doba trvání buněčného cyklu se pohybuje od několika minut do měsíců, v závislosti na typu buňky.
Buněčný cyklus je rozdělen do dvou fází, jmenovitě fáze M a rozhraní. Ve fázi M se vyskytují dva procesy, které se nazývají mitóza a cytokineze. Mitóza se skládá z jaderného dělení. Stejný počet a typy chromozomů přítomných v původním jádru se nacházejí v dceřiných jádrech. Somatické buňky mnohobuněčných organismů se dělí mitózou.
Cytokineze spočívá v rozdělení cytoplazmy za vzniku dceřiných buněk.
Rozhraní má tři fáze: 1) G1, buňky rostou a tráví většinu času v této fázi; 2) S, duplikace genomu; a 3) G2, replikace mitochondrií a dalších organel, kondenzace chromozomů a shromáždění mikrotubulů, mimo jiné..
Mitóza začíná koncem fáze G2 a je rozdělena do pěti fází: profáze, prometafáze, metafáze, anafáze a telofáze. Všichni se dějí nepřetržitě.
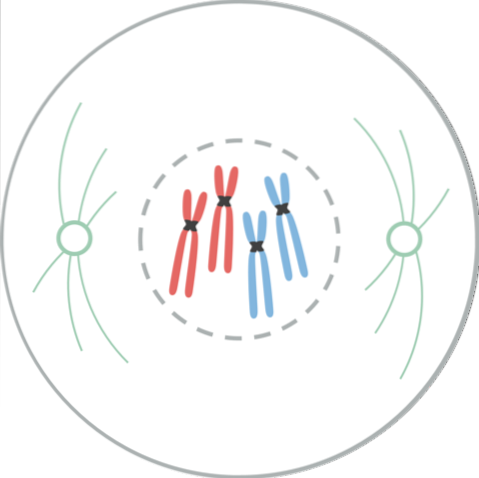
V této fázi je shromáždění mitotického vřetene nebo mitotického aparátu hlavní událostí. Profáza začíná zhutněním chromatinu za vzniku chromozomů.
Každý chromozom má sesterský chromatidový pár s identickou DNA, které jsou úzce spojeny v blízkosti jejich centromer. Na tomto spojení se účastní proteinové komplexy zvané cohesiny..
Každá centromera je připojena k kinetochoru, což je komplex proteinů, který se váže na mikrotubuly. Tyto mikrotubuly umožňují přenos každé kopie chromozomů do dceřiných buněk. Mikrotubuly vyzařují z každého konce buňky a tvoří mitotický aparát..
Ve zvířecích buňkách před profází dochází k duplikaci centrozomu, který je hlavním organizačním centrem mikrotubulů a místem, kde se setkávají rodič a dítě. Každý centrosom dosáhne opačného pólu buňky a vytvoří mezi nimi můstek mikrotubulů nazývaný mitotický aparát..
V nověji vyvinutých rostlinách na rozdíl od živočišných buněk neexistují žádné centrosomy a původ mikrotubulů je nejasný. Ve fotosyntetických buňkách staršího evolučního původu, jako jsou zelené řasy, jsou centrosomy.

Mitóza musí zajistit segregaci chromozomů a distribuci jaderného obalu komplexu jaderných pórů a nukleol. V závislosti na tom, zda jaderná obálka (EN) zmizí nebo ne, a na míře densintegrace EN, se mitóza pohybuje od uzavřené po zcela otevřenou.
Například v S. cerevisae mitóza je uzavřena, v A. nidulans je pootevřený a u lidí je otevřený.
V uzavřené mitóze se polární těla vřetena nacházejí uvnitř jaderného obalu, což tvoří nukleační body jaderných a cytoplazmatických mikrotubulů. Cytoplazmatické mikrotubuly interagují s buněčnou kůrou a s kinetochory chromozomů.
V polootevřené mitóze, protože EN je částečně rozložena, je jaderný prostor napaden nukleovanými mikrotubuly z centrosomů a dvěma otvory v EN, které tvoří svazky obklopené EN.
U otevřené mitózy dochází k úplné demontáži EN, mitotický aparát je dokončen a chromozomy se začínají přemisťovat směrem ke středu buňky.
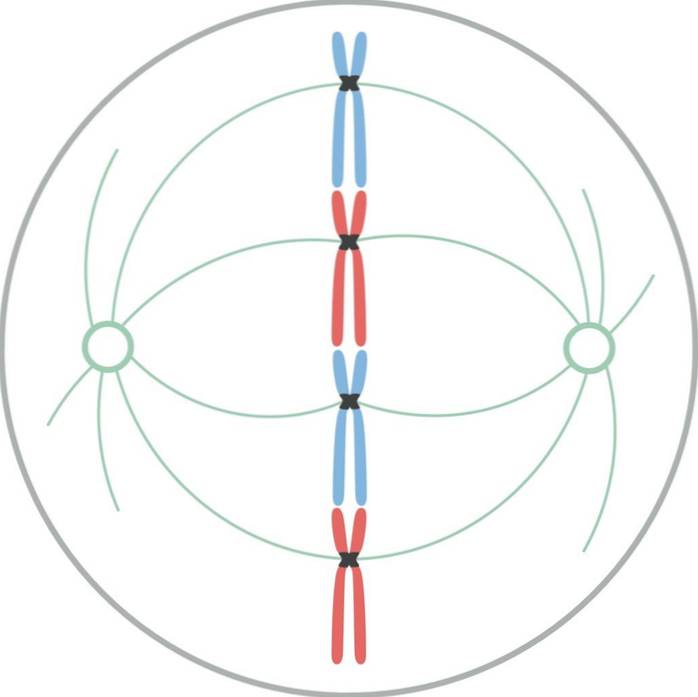
V metafázi se chromozomy seřadí na rovníku buňky. Pomyslná rovina kolmá k ose vřetena, procházející vnitřním obvodem buňky, se nazývá metafázová deska..
V savčích buňkách je mitotický aparát organizován do centrálního mitotického vřetene a dvojice asterů. Mitotické vřeteno se skládá z bilaterálního symetrického svazku mikrotubulů, který je rozdělen na rovníku buňky a tvoří dvě protilehlé poloviny. Astry jsou tvořeny skupinou mikrotubulů na každém pólu vřetena.
V mitotickém aparátu existují tři skupiny mikrotubulů: 1) astrální, které tvoří aster, vycházejí z centrosomu a vyzařují směrem k buněčné kůře; 2) kinetochore, které jsou připojeny k chromozomům přes kinetochore; a 3) polární, které se prolínají s mikrotubuly z opačného pólu.
Ve všech výše popsaných mikrotubulích jsou (-) konce obráceny k centrozomu.
Pokud v rostlinných buňkách není centrosom, je vřeteno podobné jako u živočišných buněk. Vřeteno se skládá ze dvou polovin s opačnou polaritou. Konce (+) se setkávají na rovníkové desce.
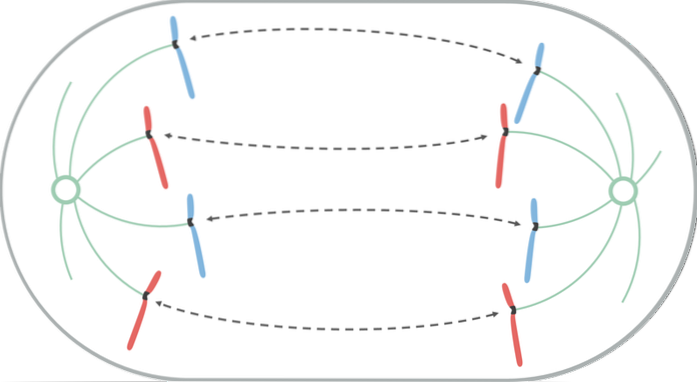
Anaphase se dělí na časnou a pozdní. K odloučení sesterských chromatidů dochází v časné anafázi.
K této separaci dochází proto, že jsou štěpeny proteiny, které udržují spojení, a protože dochází ke zkrácení mikrotubulů kinetochore. Když se dvojice sesterských chromatidů rozdělí, říká se jim chromozomy..
Během posunu chromozomů směrem k pólu se kinetochor pohybuje podél mikrotubulu stejného kinetochore, kde se jeho (+) konec disociuje. Z tohoto důvodu je pohyb chromozomů během mitózy pasivním procesem, který nevyžaduje motorické proteiny..
V pozdní anafáze dochází k většímu oddělení pólů. KRP protein, připojený k (+) konci polárních mikrotubulů, v oblasti jejich překrytí, putuje k (+) konci sousedního antiparalelního polárního mikrotubulu. KRP tedy tlačí sousední polární mikrotubule směrem k (-) konci.
V rostlinných buňkách zůstává po oddělení chromozomů uprostřed vřetena prostor s interdigitovanými nebo překrývajícími se mikrotubuly. Tato struktura umožňuje iniciaci cytokinetického aparátu, který se nazývá fragmoplast..
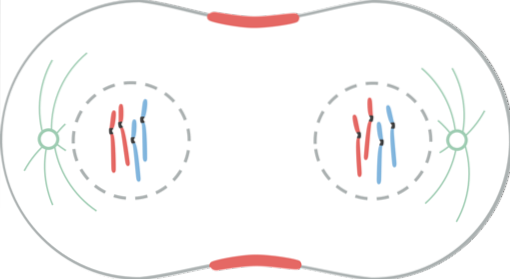
V telofázi dochází k různým událostem. Chromozomy dosahují pólů. Kinetochor zmizí. Polární mikrotubuly se nadále prodlužují a připravují buňku na cytokinezi. Jaderná obálka je přestavěna z fragmentů mateřské obálky. Nukleolus se znovu objeví. Chromozomy se stávají dekondenzovanými.
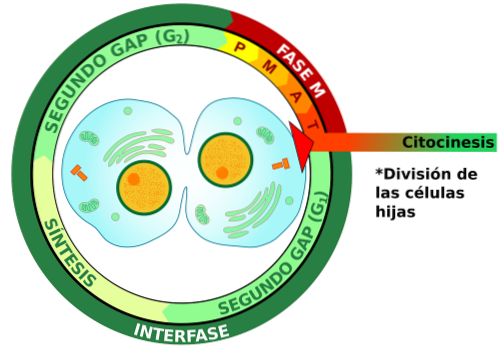
Cytokineze je fáze buněčného cyklu, během níž se buňka dělí. Ve zvířecích buňkách dochází k cytosinéze pomocí stahovacího pásu aktinových vláken. Tato vlákna klouzají kolem sebe, průměr pásky se zmenšuje a kolem obvodu buňky se tvoří štěpná drážka..
Jak zúžení pokračuje, sulcus se prohlubuje a vytváří se mezibuněčný můstek, který obsahuje střední část těla. V centrální oblasti mezibuněčného můstku jsou svazky mikrotubulů, které jsou pokryty elektrodovou matricí.
K rozpadu mezibuněčného můstku mezi postmitotickými sesterskými buňkami dochází prostřednictvím abscise. Existují tři typy abscise: 1) mechanický mechanismus rozpadu; 2) mechanismus plnění vnitřními vezikuly; 3) zúžení plazmatické membrány pro štěpení.
V rostlinných buňkách se v nich shromažďují membránové komponenty a vytváří se buněčná deska. Tento plak roste, dokud nedosáhne povrchu plazmatické membrány, spojí se s ním a rozdělí buňku na dvě části. Poté se celulóza uloží na novou plazmatickou membránu a vytvoří novou buněčnou stěnu..
Meióza je typ buněčného dělení, který snižuje počet chromozomů na polovinu. Diploidní buňka se tedy dělí na čtyři haploidní dceřiné buňky. Meióza se vyskytuje v zárodečných buňkách a vede ke vzniku gamet.
Stádia meiózy se skládají ze dvou rozdělení jádra a cytoplazmy, jmenovitě meiózy I a meiózy II. Během meiózy I se členové každé dvojice homologních chromozomů oddělují. Během meiózy II se sesterské chromatidy oddělují a produkují se čtyři haploidní buňky.
Každá fáze mitózy je rozdělena na profázu, prometafázu, metafázi, anafázi a telofázu..
- Prophase I. Chromozomy kondenzují a vřeteno se začíná formovat. DNA byla duplikována. Každý chromozom je tvořen sesterskými chromatidy připojenými k centromere. Homologní chromozomy se během synapse spárují, což umožňuje křížení, což je klíč k produkci různých gamet.
- Metafáze I. Dvojice homologních chromozomů se seřadí podél metafázové desky. Chiasma pomáhá držet pár pohromadě. Mikrotubuly kinetochore na každém pólu se vážou na centromeru homologního chromozomu.
- Anafáze I. Mikrotubuly kinetochore jsou zkráceny a homologické páry jsou odděleny. Jeden duplikát homolog jde na jeden pól buňky, zatímco druhý duplikát homolog jde na druhou stranu pólu..
- Telophase I. Samostatné homology tvoří skupinu na každém pólu buňky. Jaderná obálka se znovu formuje. Cytokineze se děje. Výsledné buňky mají poloviční počet chromozomů než původní buňky.
- Prophase II. V každé buňce se vytvoří nové vřeteno a buněčná membrána zmizí.
- Metafáze II. Vytváření vřetena je dokončeno. Chromozomy mají sesterské chromatidy, spojené v centromere, zarovnané podél metafázové desky. Mikrotubuly kinetochore, které začínají od protilehlých pólů, se vážou na centromery.
- Anafáze II. Mikrotubuly se zkracují, centromery se dělí, sesterské chromatidy se oddělují a pohybují se směrem k opačným pólům.
- Telophase II. Jaderný obal je tvořen kolem čtyř skupin chromozomů: jsou tvořeny čtyři haploidní buňky.
Některé příklady ilustrují důležitost různých typů buněčného dělení.
- Mitóza Buněčný cyklus má nevratné body (replikace DNA, separace sesterských chromatid) a kontrolní body (G1 / S). Protein p53 je klíčem ke kontrolnímu bodu G1. Tento protein detekuje poškození DNA, zastavuje dělení buněk a stimuluje aktivitu enzymů, které poškození opravují..
U více než 50% lidských rakovin má protein p53 mutace, které ruší jeho schopnost vázat specifické sekvence DNA. Mutace v p53 mohou být způsobeny karcinogeny, jako je benzopyren v cigaretovém kouři.
- Redukční dělení buněk Je spojena se sexuální reprodukcí. Z evolučního hlediska se předpokládá, že sexuální reprodukce vznikla jako proces opravy DNA. Poškození chromozomu lze tedy opravit na základě informací z homologního chromozomu..
Předpokládá se, že diploidní stav byl ve starodávných organismech přechodný, ale s přibývajícím genomem se stal relevantnějším. V těchto organismech má pohlavní rozmnožování funkci komplementace, opravy DNA a genetické variace..



Zatím žádné komentáře