
The postmendelské dědictví Zahrnuje všechny myšlenky a / nebo práce, které vznikly po studiích publikovaných náboženským přírodovědcem Gregoriem Mendelem v roce 1865 a které vysvětlují genetické jevy nebo chování odlišné od těch, které popisuje.
Gregorio Mendel je považován za otce genetiky, protože studiem křížení rostlin hrachu vytvořil první zákony dědičnosti před více než 100 lety. První zákon, nazvaný „Zákon segregace“, navrhuje, aby geny mohly existovat v různých nebo alternativních formách.

Tyto alternativní formy se nazývají „alely“ a přímo ovlivňují fenotypové vlastnosti druhu. Alely mohou určit barvu vlasů, barvu očí, typ vlasů atd..
Mendel ve svém prvním zákoně navrhl, aby každý fenotypový znak přenášený z rodičů na jejich děti byl řízen dvěma alelami, každá pocházející od rodiče (jedna od „matky“ a jedna od „otce“).
Podle toho, když rodiče přispívají různými alelami, vede potomstvo ke směsi, kde jedna z alel je dominantní (ta, která je vyjádřena ve fenotypu) a druhá je recesivní (není vidět, nebo je „latentní“ ")..
Mendelovo druhé právo je známé jako „Zákon nezávislého dědictví“, protože Mendel ve svých kříženích poznamenal, že vzor dědičnosti, který charakterizuje alely, které definují znak, neovlivňuje vzor dědičnosti alel, které definují jiný znak..
Rejstřík článků
Od doby, kdy Gregor Mendel navrhl své slavné zákony před více než 100 lety, byly objasněny nové mechanismy vysvětlující dědičnost postav, z nichž mnohé lze považovat za protikladné nebo značně odlišné od těch, které popsal Mendel..
Příkladem jevů popsaných „moderní“ vědou, jejichž pokroky byly možné pouze díky pokroku v experimentálních strategiích a výzkumných metodách, jsou mimo jiné dědičnost spojená se sexem, neúplná dominance a kodominance..
Dědičnost spojená se sexem souvisí s geny, které se nacházejí pouze na pohlavních chromozomech. U lidí je pohlaví určováno chromozomy X a Y. Ženy mají dvě kopie X (XX) a muži mají jednu z X a jednu z Y (XY)..
Geny nalezené na každém z těchto dvou chromozomů jsou ty, které přispívají k těm vlastnostem, které jsou spojeny se sexem. Geny na chromozomu X jsou přítomny u mužů a žen, ale geny Y jsou pouze u mužů.
Dědičnost spojená se sexem byla poprvé pozorována a popsána Williamem Batesonem, jedním z prvních vědců, kteří „znovuobjevili“ Mendelovu práci a využili ji k provedení nových experimentů na základě jeho závěrů..
Bateson vyrobil experimentální kříže slepic s peřím různých barev, některé s tmavým a hladkým peřím a jiné s hladkým peřím a pruhovaným vzorem..
Prostřednictvím těchto křížů si všiml, že u potomků měli výslední muži vždy peří s pruhovaným vzorem.
Teprve ve třetí generaci, kdy byl pruhovaný samec zkřížen se slepicí s tmavým a hladkým peřím, byli získáni někteří kohouti s černým peřím. Tento výsledný podíl křížů vzdoroval proporcím navrženým Mendelem.
Po Batesonově díle našel Thomas Henry Morgan logické vysvětlení podivných výsledků, které Bateson získal, připisující rozdíl v zabarvení kuřat skutečnosti, že pruhovaný vzor byl kódován dominantní alelou nalezenou na X chromozomu..
U slepic, na rozdíl od lidí, odpovídá heterogametické pohlaví (XY) ženám, zatímco homogametické pohlaví odpovídá mužům (XX).
Alelu studovanou Batesonem a Morganem pak vyjádřili všichni muži s pruhovaným opeřením, ale když muž zdědil dva chromozomy X s alelou odpovídající černé barvě, objevili se muži s tmavým opeřením a bez pruhovaného vzoru..
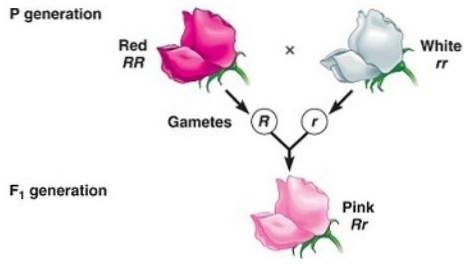
Neúplná dominance je, když fenotyp pozorovaný u potomků, který je výsledkem křížení dvou homozygotních jedinců (jednoho dominantního a druhého recesivního), je „přechodným“ fenotypem mezi dominantním a recesivním..
Souvisí to s „neschopností“ „dominantní“ alely úplně maskovat expresi recesivní alely, a proto výsledný fenotyp odpovídá směsi znaků kódovaných oběma alelami, mnohokrát popsaných jako „zředěný“ fenotyp “Dominantní.
Například v závodech Dondiego v noci (Mirabilis jalapa) typická barva okvětních lístků divoké květiny je červená. Když je rostlina s červenými okvětními lístky zkřížena s rostlinou čisté linie s bílými okvětními lístky, získá se, že všichni potomci jsou růžové (růžové) okvětní lístky.
Když jsou zkříženi dva dominantní homozygotní jedinci a u potomků je pozorován třetí fenotyp, který odpovídá dvěma charakteristikám určeným dvěma rodičovskými alelami, hovoříme o kodominanci.
V tomto případě, pokračujícím příkladem podobným jako u květů v předchozím příkladu, by „třetí fenotyp“ neodpovídal růžovým květům, ale rostlině s bílými květy a červenými květy na stejném stonku, protože tyto dvě alely „codominate“, a ani jeden maskuje toho druhého.
Mnohokrát je však obtížné fenotypicky odlišit kodominanci a neúplnou dominanci, protože obě produkují fenotyp jedince, který, jak se zdá, vyjadřuje dvě různé alely..
Genotypicky se však jedinci, kteří představují alely kodominantní, liší od těch, kteří vykazují neúplnou dominanci, protože v jednom případě jsou dvě dominantní alely vyjádřeny ve stejném množství a v druhém jsou podíly nerovné..
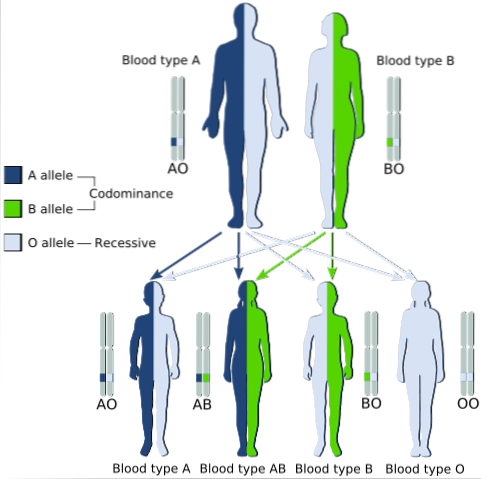
Klasickým příkladem kodominance je typ krevní skupiny u lidí, protože alely kódující antigeny A a B v lidské populaci jsou kodominantní.
To znamená, že když se alely A a B nacházejí společně v buňkách jednotlivce, má krevní skupinu AB.
Jsou to recesivní alely pro nějakou postavu, které, pokud jsou nalezeny v homozygotní formě, jsou pro jednotlivce smrtelné. Jedinci, kteří tyto alely prezentují, se nenarodili, protože brání normálnímu fungování molekulárního aparátu a brání translaci genů.
Mnoho míst v genomu jedince interaguje s jinými částmi stejného genomu, to znamená, že specifická modifikace alely v genomu může přímo podmínit expresi dalších alel v buňkách jedince.
Tyto interakce je obtížné určit a jsou klasifikovány do dvou různých typů: geny, které interagují ve stejné genetické dráze a ty, které interagují v různých genetických drahách..
Klasický a známý příklad genů, které interagují různými genetickými cestami, souvisí s geny, které určují barvu kůže kukuřičných hadů (Pantherophis guttatus), jehož společnou barvou je opakující se vzor černých a oranžových skvrn.

Tento fenotyp je produkován dvěma různými pigmenty, které jsou pod přísnou genetickou kontrolou. Gen určuje oranžovou barvu a má dvě alely: jednu O +, která kóduje pigment, a druhou O-, která určuje celkovou nepřítomnost oranžového pigmentu..
Další odlišný gen určuje přítomnost nebo nepřítomnost černého pigmentu, alely tohoto genu jsou známé jako B +, ten, který určuje přítomnost černého pigmentu a B-, který určuje nepřítomnost pigmentu.
Oba geny nejsou spojeny a nezávisle určují barvu hadů, to znamená, že had s genotypem O + / B + je normální had s oranžovým a černým fenotypem, ale jakákoli genotypová variace bude mít za následek jiný fenotyp..
Klasickým příkladem těchto genů je gen popsaný pro „zvonkovité“ květiny (čeledi Campanulaceae), jejichž „divoký“ fenotyp odpovídá rostlinám s modrými květy..
Modrá barva květů zvonků není určena genem, ale metabolickou cestou, kde se na syntéze pigmentu podílí několik enzymů, každý kódovaných jiným genem..
Existují tedy fenotypy charakterizované tím, že jsou výsledkem interakce několika genů, jejichž produkty se účastní stejné metabolické dráhy a kde mutace nebo nesprávná funkce kteréhokoli z nich vede k fenotypu, v tomto případě bílých, depigmentovaných květů..

Zatím žádné komentáře