The lišejníky jsou symbiotickými asociacemi mezi houbou (mykobiont) a zelenou řasou nebo sinicemi (fotobiont). Plísně tvořící houby nemohou v přírodě přežít samy a bez jejich fotobiontu nemohou generovat velkou rozmanitost růstových forem lišejníků nebo sekundárních látek..
Většina mykobiontů patří do skupiny Ascomycota zvané Lecanoromycetes. Většina fotobiontů patří k rodům Trebouxia Y Trentepohlia (zelené řasy) a Calothrix, Gloecapsa Y Nostoc (sinice).

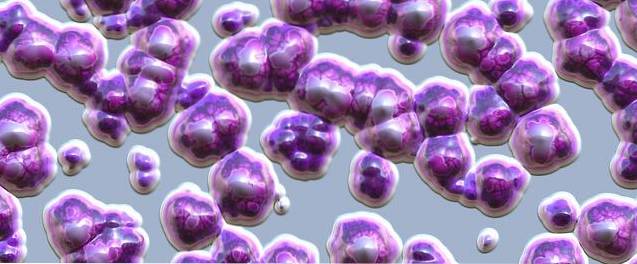
Na první pohled vypadají lišejníky jako rostliny, ale prostřednictvím mikroskopu vidíme asociaci milionů fotobiontových buněk propletených v matrici tvořené vlákny houby. Houba tvoří thallus, ve kterém je umístěn fotobiont.
Asi 8% suchozemských ekosystémů ovládají lišejníky. V těchto ekosystémech jsou cévnaté rostliny na své fyziologické hranici. Lišejníky mají výhodu v jejich schopnosti přežít extrémní chlad, teplo a vodní stres, a proto mohou zůstat ve stavu letargie.
Lišejníky se vyznačují jejich distribucí, množením a množením, morfologií, metabolismem, symbiotickými interakcemi a ekologií..
Rejstřík článků
Lišejníky se vyskytují téměř všude na světě, zejména v extrémním prostředí, jako je poušť a vysoké hory. Existuje úzký vztah mezi tvarem thallus (také nazývaným tělo lišejníku) a jeho distribucí. Thallus má tři různé růstové formy: crustose, foliose a fruktózu.
Crustose thallus připomíná kůru úzce spojenou s povrchem. Nelze je odstranit, aniž by došlo ke zničení lišejníků. Lišejníky tohoto tvaru odolávají suchu a jsou dobře přizpůsobeny suchému podnebí, jako je poušť. Příkladem je Arthopyrenia halodyty který žije ve Středozemním moři na vápenatých substrátech.
Listový (nebo listový) thallus připomíná malý keř. Lišejníky tohoto tvaru rostou nejlépe v oblastech s častým deštěm. Příkladem je pohlaví Physma, který žije v tropickém deštném pralese v Austrálii, na kůře stromů.
Fruticus (nebo fruticulous) thallus je vláknitý, ve tvaru listu. Lišejníky tohoto tvaru používají atmosférickou vodní páru. Žijí hlavně ve vlhkém prostředí, jako jsou zatažené oblasti na pobřeží oceánu a horské oblasti v tropech. Příkladem je Kuřecí větev který žije na jedleAbies alba) ve Švýcarsku.
Nejběžnější reprodukcí lišejníků je sexuální mykobiont. U tohoto typu reprodukce mykobiont uvolňuje četné spory, které po klíčení musí najít kompatibilní fotobiont.
Vzhledem k tomu, že spory jsou geneticky rozmanité, spojení houby a zelených řas za vzniku lišejníků vytváří u lišejníků velkou genetickou variabilitu. Je třeba poznamenat, že fotobiont se reprodukuje pouze klonálně, s výjimkou fotobiontů, které patří k Trentepohliales.
Pokud se mykobiont reprodukuje nepohlavně, přenáší se fotobiont se svou mykobiontem na další generaci prostřednictvím specializovaných vegetativních propagul, jako jsou soredia a isidia. Jedná se o vnější výrůstky prasklinami a póry na povrchu kůry thallus..
Soredia jsou malé shluky buněk řas a plísňových mycelií. Tento způsob šíření je typický pro lišejníky foliosy a plody. Například thallus z Lepraria sestává zcela ze soredios.
Isidia jsou malá rozšíření thallus, která také slouží k nepohlavnímu šíření, pokud jsou odříznuta od thallus. Například thallus z Parmotrema crinitum je pokryta isidií.
Morfologie a anatomie lišejníků odpovídá omezením, která na symbiózu ukládá životní prostředí. Mycobiont je vnější a fotobiont vnitřní. Vzhled thallusu určuje mykobiont.
Všechny lišejníky mají podobnou vnitřní morfologii. Tělo lišejníku je tvořeno vlákny mykobiontu.
Hustota těchto vláken definuje vrstvy lišejníku. Na povrchu, který je ve styku s prostředím, jsou vlákna velmi zhutněna a tvoří kůru, což snižuje intenzitu světla a brání poškození fotobiontu..
Pod kůrou je vrstva tvořená řasami. Tam je hustota vláken nízká. Pod vrstvou řas je dřeň, což je volná vrstva složená z vláken. U lišejníků crustose se dřeň dotýká substrátu.
V lišejníkových lišejnících je pod dřeně druhá kůra, nazývaná vnitřní kůra, která je k substrátu připevněna hyfy houby, které se podobají kořenům, a proto se jim říká rhiziny.
U ovocných lišejníků kůra obklopuje vrstvu řas. To zase obklopuje dřeň.
Asi 10% z celkové biomasy lišejníků je tvořeno fotobiontem, který syntetizuje sacharidy fotosyntézou. Mezi 40% a 50% suché hmotnosti lišejníků je uhlík fixovaný fotosyntézou.
Sacharidy syntetizované ve fotobiontu jsou transportovány do mykobiontu, kde jsou použity pro biosyntézu sekundárních metabolitů. Pokud je fotobiontem sinice, syntetizovaným sacharidem je glukóza. Pokud se jedná o zelené řasy, jsou uhlohydráty ribitol, erytrol nebo sorbitol.
Hlavní třídy sekundárních metabolitů pocházejí z:
- Acetyl-polymalonyl
- Kyselina mevalonová
- Kyselina shikimová.
Produkty první cesty jsou alifatické kyseliny, estery a příbuzné deriváty, jakož i aromatické sloučeniny odvozené od polyketidů. Produkty druhé cesty jsou triterpeny a steroidy. Produkty třetí cesty jsou terfenylchinony a deriváty kyseliny pulvinové.
Fotobiont také dodává mykobiontu vitamíny. Mycobiont mu sám poskytuje vodu získanou ze vzduchu a vystavuje fotobiont světlu, aby mohl provádět fotosyntézu. Pigmenty nebo krystaly přítomné v kůře působí jako filtry a absorbují určité vlnové délky nezbytné pro fotosyntézu..
Pro symbiotické asociace lze použít pojmy selektivita a specificita. Selektivita je, když jeden organismus přednostně interaguje s druhým. Specifičnost se týká interakce buňka-buňka, ve které existuje absolutní exkluzivita.
Bylo navrženo, že lišejníky lze považovat za vysoce selektivní symbiózu. Některá pozorování, která podporují tuto myšlenku, jsou:
- Z tisíců rodů řas je velmi málo fotobiontů.
- Určité volné řasy, které kolonizují stejná stanoviště, lišejníky do nich nejsou začleněny, přestože jsou v přímém kontaktu.
Bylo navrženo, že u některých lišejníků, jako jsou rody Cladonia, existuje silná selektivita a specificita mykobiontu vůči symbiontové řase. Jiné lišejníky, například rody Lepraria Y Stereocaulon vykazují pouze specifičnost (v obou případech vůči řasám) Asterochloris).
Obecně je specificita na úrovni druhu nebo populace nízká. Dále je třeba vzít v úvahu, že specificita není jediným determinantem složení: asociace mezi jednotlivci je ovlivněna místními podmínkami prostředí..
Ve srovnání s cévnatými rostlinami jsou lišejníky špatnou konkurencí kvůli své malé velikosti a extrémně pomalému růstu. Přes toto může složení lišejníků ovlivnit strukturu a chemii půdy, zvýšit pokrytí a biodiverzitu..
Přítomnost a hojnost lišejníků je dána faktory, jako je chemie a stabilita substrátu, dostupnost světla a vlhkost prostředí. Společenstva lišejníků se tedy mohou měnit v důsledku teploty nebo dostupnosti vody..
Z tohoto důvodu lišejníky slouží jako bioindikátory změny klimatu, které lze pravidelně sledovat analýzou pokrytí a druhové bohatosti lišejníků přítomných ve studované oblasti..
Používání lišejníků jako bioindikátorů změny klimatu má následující výhody:
- Není nutné denní měření.
- Lišejníky mají dlouhou životnost a jsou široce distribuovány.
- Monitorování lišejníků lze provádět na stanicích v regionech s extrémními podmínkami prostředí.
Fotobionty některých lišejníků také slouží jako bioindikátory znečištění životního prostředí. Například fotobiont Coccomyxa je velmi citlivý na těžké kovy.
Lišejníky vykazují výraznou odolnost a jsou schopné se usadit v nehostinném prostředí pro jiné živé bytosti. Mohou však být také velmi citliví na narušení životního prostředí způsobené lidmi..
Lišejníky lze klasifikovat podle prostředí, ve kterém rostou, podle jejich požadavků na pH nebo podle typu živin, které odebírají ze substrátu. Například podle prostředí se lišejníky dělí na saxikoly, kortikosteroidy, mořské, sladkovodní a folikuly..
Saxikulturní lišejníky rostou na skalách. Příklad: Mučivý film, Amandinea coniops, Verrucaria elaeina.
Na kůře stromů rostou zahradnické lišejníky. Příklady: Alectoria spp., Cryptothecia rubrocincta, Evernia spp., Lobaria pulmonaria, Usnea spp.
Mořské lišejníky rostou na skalách, kde vlny bijí. Příklady: Arthopyrenia halodyty, Liči spp., Verrucaria maura.
Sladkovodní lišejníky rostou na skalách, na kterých se pohybuje voda. Příklady: Peltigera hydrothyria, Leptosira obovata.
Folikulární lišejníky rostou na listech deštného pralesa. Druhy tohoto typu slouží jako mikroklimatické bioindikátory.
Protože jsou to polyspecifické organismy a jsou považovány za součet mykobiontů a mykobiontů, lišejníky postrádají formální status v taxonomii živých organismů. Starověké taxonomické klasifikace lišejníků jako jednotlivých entit se vyvinuly dříve, než byla uznána jejich symbiotická povaha.
Současná taxonomie lišejníků je založena výhradně na charakterech a fylogenetických vztazích mykobionta. Proto jsou všechny lišejníky klasifikovány jako houby..
V současné době jsou řády, rodiny a rody lišejníkovitých hub vymezeny znaky plodnic. Lišejníky s thalluses, i když jsou morfologicky odlišné, zůstávají sjednoceny ve stejné rodině nebo rodu. Uvažuje se také o jiných strukturách, jako jsou isidiany a sorediany.
98% druhů hub, které tvoří lišejníky, patří do kmene Ascomycota. Většina zbývajících druhů patří do kmene Basidiomycota. Pokud jde o fotobionty, 87% druhů tvoří zelené řasy, 10% sinice a 3% kombinace zelených řas a sinic..
Molekulární studie umožnily upravit koncept druhů na základě morfologie. Podobně studie sekundárních metabolitů umožnily oddělení morfologicky podobných druhů..
Protože lišejníky jsou primárními producenty, slouží jako potrava pro býložravá zvířata. V Severní Americe a Eurasii se velké býložravé savce, jako jsou sobi a karibu, živí lišejníky Cladonia rangiferina. V zimě mohou tito býložravci jíst tento lišejník mezi 3 a 5 kg denně.
C. rangiferina, známý jako lišejník sobů, patří do třídy Lecanoromycetes a do čeledi Cladoniaceae. C. rangifera může dosáhnout velikosti podobné jako u typických cévnatých rostlin. Má šedou barvu a ovocný talus.
Druhy patřící do rodu Cladonia jsou tolerantní k vysokým koncentracím kovů, proto mohou uchovávat vysoké koncentrace radioaktivních derivátů stroncia a cesia. Spotřeba tohoto lišejníku zvířaty představuje problém, protože u mužů, kteří tato zvířata jedí, může dosáhnout škodlivých hladin.
Evernia prunastri, známý jako dubový mech, a Pseudevergne furfuracea, Známý jako stromový mech, jsou důležitými druhy lišejníků v parfémovém průmyslu. Patří do třídy Lecanoromycetes a do čeledi Parmeliaceae.
Oba druhy se shromažďují na jihu Francie, Maroka a bývalé Jugoslávie a ročně se zpracovává přibližně 9 000 tun. Kromě toho, že jsou užitečné pro parfémový průmysl, P. furfuracea je citlivý na znečištění, a proto se používá ke sledování průmyslové kontaminace.
Lišejníky jsou bohaté na pigmenty, které slouží k blokování ultrafialového B (UVB) světla. Lišejníkové sinice Collema je bohatý na tento typ pigmentů, které byly čištěny a patentovány jako produkt, který poskytuje 80% ochranu proti UVB.
Kyanoliquen Collema cristatum, například máte pigment zvaný collemin A (ʎmax= 311 nm), což je mykosporin, který poskytuje ochranu proti UVB záření (280 - 315 nm).
Roccellla montagnei Jedná se o ovocný likér, který roste na skalách, ze kterého se ve středomořské oblasti získává červené nebo fialové barvivo. Jiné lišejníky jako Heteroderma obscurata Y Nephroma laevigatum obsahují antrachinony používané jako barviva.
Lišejníky obsahují látky, které by mohly být použity ve farmaceutickém průmyslu. Mnoho druhů lišejníků má aktivní sloučeniny, které ničí bakterie, jako jsou Zlatý stafylokok, Pseudomonas aeruginosa, Bacillus subtilis Y Escherichia coli. Kromě toho mají lišejníky vysoký potenciál jako zdroj protinádorových léků..


Zatím žádné komentáře