The mikrofilamenta nebo aktinová vlákna, jsou jednou ze tří základních složek cytoskeletu eukaryotických buněk (mikrofilamenta, mikrotubuly a mezivlákna) a jsou složena z malých vláken proteinu zvaného aktin (aktinové polymery).
U eukaryot jsou geny kódující aktinová mikrofilamenta vysoce konzervované ve všech organismech, a proto se často používají jako molekulární markery pro studium různých druhů.

Mikrovlákna jsou distribuována po celém cytosolu, ale hojně se vyskytují zejména v oblasti pod plazmatickou membránou, kde tvoří složitou síť a spojují se s dalšími speciálními proteiny za vzniku cytoskeletu..
Sítě mikrofilament v cytoplazmě savčích buněk jsou kódovány dvěma ze šesti genů popsaných pro aktin, které se účastní dynamiky mikrofilamentů a jsou dokonce velmi důležité během diferenciace kmenových buněk.
Mnoho autorů souhlasí s tím, že mikrofilamenta jsou nejrozmanitější, nejuniverzálnější a nejdůležitější proteiny v cytoskeletu většiny eukaryotických buněk, a je důležité si uvědomit, že se v prokaryotických mikroorganismech nenacházejí..
Na druhé straně v tomto typu buněk existují vlákna, která jsou homologní s mikrofilamenty, ale která jsou vytvořena z jiného proteinu: proteinu MreB..
V současné době je gen kódující tento protein považován za možný gen předků pro eukaryotický aktin. Sekvenční homologie aminokyselin, které tvoří protein MreB, je však pouze 15% vzhledem k sekvenci aktinu.
Jelikož jsou základní součástí cytoskeletu, může jakýkoli fenotypový defekt v mikrotubulích a intermediárních vláknech a aktinových mikrofilamentách (cytoskelet) způsobit různé buněčné a systémové patologie.
Rejstřík článků
Mikrovlákna jsou tvořena proteinovými monomery rodiny aktinů, což jsou vysoce hojné kontraktilní proteiny v eukaryotických buňkách, protože se také účastní svalové kontrakce..
Tato vlákna mají průměr mezi 5 a 7 nm, takže jsou také známá jako tenká vlákna a jsou složena ze dvou forem aktinu: globulární formy (G aktin) a vláknité formy (F aktin)..
Proteiny, které se účastní cytoskeletu, jsou známé jako y a β aktiny, zatímco ty, které se účastní kontrakce, jsou obvykle α aktiny..
Podíl globulárního aktinu a vláknitého aktinu v cytosolu závisí na potřebách buněk, protože mikrofilamenta jsou vysoce variabilní a univerzální struktury, které neustále rostou a zkracují se polymerací a depolymerací..
G aktin je malý globulární protein, složený z téměř 400 aminokyselin a s molekulovou hmotností přibližně 43 kDa.
G-aktinové monomery, které tvoří mikrofilamenta, jsou uspořádány ve formě spirálového vlákna, protože každý podléhá zkroucení, když je spojen s dalším.
G aktin se asociuje s jednou molekulou Ca2 + a druhou s ATP, které stabilizují její globulární formu; zatímco F-aktin se získává po hydrolýze terminálního fosfátu molekuly ATP na G-aktin, což přispívá k polymeraci.
Aktinová vlákna lze uspořádat do „svazků“ nebo „sítí“, které mají v buňkách různé funkce. Svazky tvoří paralelní struktury spojené poměrně tuhými příčnými mosty.
Sítě jsou naproti tomu volnější struktury, jako trojrozměrné sítě s vlastnostmi polotuhých gelů..
Existuje mnoho proteinů, které jsou spojeny s aktinovými vlákny nebo mikrofilamenty a jsou známé jako ABP (z angličtiny proteiny vázající aktin), které mají konkrétní weby.
Mnoho z těchto proteinů umožňuje mikrofilamentům interagovat s dalšími dvěma složkami cytoskeletu: mikrotubuly a mezivlákny, stejně jako s dalšími složkami vnitřní strany plazmatické membrány..
Mezi další proteiny, se kterými mikrofilamenta interagují, patří jaderné vrstvy a spektrin (v červených krvinkách).
Jelikož se globulární aktinové monomery vážou vždy stejným způsobem, orientované stejným směrem, mají mikrofilamenty definovanou polaritu se dvěma konci: jedním „více" a jedním „méně"..
Polarita těchto vláken je velmi důležitá, protože rostou podstatně rychleji na svém pozitivním konci, kde jsou přidávány nové monomery G-aktinu..

První věcí, která probíhá během polymerace aktinových vláken, je proces známý jako „nukleace“, který spočívá ve spojení tří monomerů proteinu.
K tomuto trimeru se na obou koncích přidávají nové monomery, aby vlákno rostlo. Monomery G-aktinu jsou schopné hydrolyzovat ATP s každou vazbou, což má důsledky pro rychlost polymerace, protože skupiny aktin-ATP disociují s většími obtížemi než skupiny aktin-ADP..
ATP není pro polymeraci nezbytný a specifická role jeho hydrolýzy dosud nebyla objasněna..
Někteří autoři se domnívají, že jelikož události polymerace aktinů jsou rychle reverzibilní, může ATP spojená s těmito procesy představovat až 40% celkového buněčného obratu této energetické molekuly.
Jak polymerace aktinových vláken, tak jejich depolymerace jsou procesy vysoce regulované řadou specifických proteinů, které jsou odpovědné za remodelaci vláken..
Příkladem proteinů, které regulují depolymeraci, je aktin depolymerizační faktor cofilin. Jiný protein, profilin, má opačnou funkci, protože stimuluje asociaci monomerů (stimulací výměny ADP za ATP).
Mikrovlákna interagují s myosinovými vlákny, která jsou spojena s transmembránovými proteiny, které mají doménu v cytosolu a jinou v buněčném zevnějšku, čímž se účastní procesů buněčné mobility.
Tato mikrofilamenta spojená s plazmatickou membránou zprostředkovávají různé buněčné reakce na různé třídy stimulů. Například buněčná adheze v epiteliálních tkáních je řízena transmembránovými proteiny známými jako kadheriny, které interagují s mikrofilamenty a získávají faktory odezvy..
Aktinová vlákna interagují s intermediárními vlákny a způsobují přenos extracelulárních stimulů do klíčových míst, jako jsou ribozomy a chromozomy v jádru..
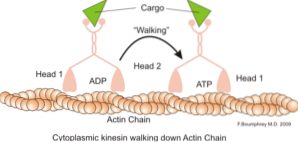
Klasickou a hodně studovanou funkcí mikrofilament je jejich schopnost tvořit „mosty“, „kolejnice“ nebo „dálnice“ pro pohyb motorického proteinu myosin I, který je schopen v sekreční sekvenci přenášet vezikuly z organel na membránový plazmatický cesty.
Mikrovlákna také interagují s myosinem II a vytvářejí kontraktilní kruh, který se tvoří během cytokineze, přesně během posledního stadia buněčného dělení, ve kterém je cytosol oddělen od kmenových a dceřiných buněk..
Obecně F-aktinová mikrofilamenta modulují distribuci některých organel, jako je Golgiho komplex, endoplazmatické retikulum a mitochondrie. Kromě toho se také podílejí na prostorovém umístění mRNA, takže jsou čteny ribozomy..
Celá buněčná sada mikrofilament, zejména těch, která úzce souvisí s plazmatickou membránou, se podílí na tvorbě zvlněných membrán buněk, které mají stálý aktivní pohyb.
Podílejí se také na tvorbě mikroklků a dalších běžných hrbolů na povrchu mnoha buněk..
Mikrovlákna se účastní procesu sekrece žluči v hepatocytech (jaterní buňky) a také na peristaltických pohybech (koordinovaná kontrakce) jaterních kanálků.
Přispívají k diferenciaci domén plazmatické membrány díky jejich asociaci s různými cytosolickými elementy a kontrole nad topografií těchto intracelulárních elementů..
Existuje několik onemocnění spojených s primárními defekty ve struktuře nebo s regulačními proteiny a enzymy při syntéze mikrofilament, a to navzdory skutečnosti, že se přímo podílejí na velkém počtu funkcí..
Nízká míra onemocnění a malformací v primární struktuře mikrofilament je způsobena skutečností, že obecně existuje více genů kódujících jak aktin, tak jeho regulační proteiny, což je jev známý jako „genetická redundance“.
Jednou z nejvíce studovaných patologií je vitrifikace oocytů na jejich cytoskeletu, kde je pozorováno přerušení sítě kortikálních mikrofilament, stejně jako depolymerace a dezorganizace mikrotubulů mitotického vřetena..
Obecně tato vitrifikace způsobuje chromozomální disperzi, protože vede k narušení při zhutňování veškerého chromatinu.
Buňky, které mají ve svém cytoskeletu větší organizaci a podíl mikrofilament, jsou buňkami příčně pruhovaného svalu, proto je většina patologií spojena s poruchou kontraktilního aparátu.
Defektní nebo atypická mikrofilamenta byla také spojována s onemocněním kostí známým jako Pagetova choroba..


Zatím žádné komentáře