The RNA nebo RNA (ribonukleová kyselina) je typ nukleové kyseliny přítomné v eukaryotických a prokaryotických organismech a ve virech. Jedná se o nukleotidový polymer, který ve své struktuře obsahuje čtyři typy dusíkatých bází: adenin, guanin, cytosin a uracil.
RNA se obecně vyskytuje jako jeden pás (s výjimkou některých virů), lineárně nebo v řadě složitých struktur. Ve skutečnosti má RNA strukturální dynamiku, která není pozorována v dvojité šroubovici DNA. Různé typy RNA mají velmi rozmanité funkce.
Ribozomální RNA jsou součástí ribozomů, struktur odpovědných za syntézu proteinů v buňkách. Messenger RNA fungují jako prostředníci a transportují genetickou informaci do ribozomu, který překládá zprávu z nukleotidové sekvence na aminokyselinovou sekvenci..
Transfer RNA jsou odpovědné za aktivaci a přenos různých typů aminokyselin - celkem 20 - do ribozomů. Pro každou aminokyselinu existuje molekula přenosové RNA, která rozpoznává sekvenci v messengerové RNA.
Kromě toho existují další typy RNA, které se přímo nepodílejí na syntéze proteinů a nepodílejí se na regulaci genů..
Rejstřík článků
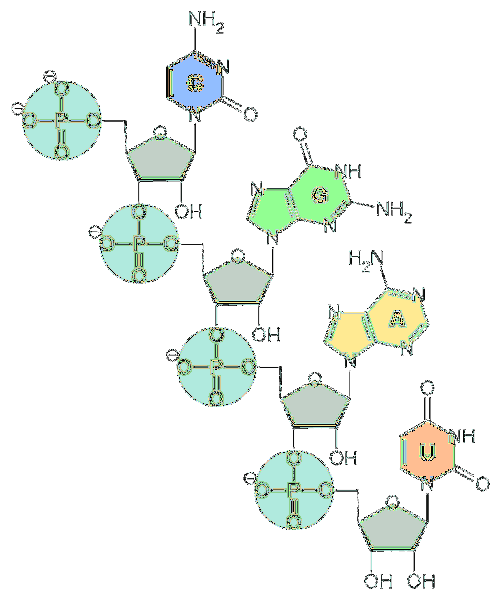
Základní jednotkou RNA jsou nukleotidy. Každý nukleotid je tvořen dusíkatou bází (adenin, guanin, cytosin a uracil), pentózou a fosfátovou skupinou..
Dusíkaté báze jsou odvozeny ze dvou základních sloučenin: pyrimidinů a purinů..
Bázemi odvozenými od purinů jsou adenin a guanin a bázemi odvozenými od pyrimidinů jsou cytosin a uracil. I když se jedná o nejběžnější báze, nukleové kyseliny mohou mít i jiné typy bází, které jsou méně časté..
Pokud jde o pentózu, jsou to jednotky d-ribózy. Z tohoto důvodu se nukleotidy, které tvoří RNA, nazývají „ribonukleotidy“.
Nukleotidy jsou navzájem spojeny chemickými vazbami zahrnujícími fosfátovou skupinu. K jejich vytvoření je fosfátová skupina na 5 'konci nukleotidu připojena k hydroxylové skupině (-OH) na 3' konci dalšího nukleotidu, čímž se vytvoří vazba podobná fosfodiesteru..
Podél řetězce nukleové kyseliny mají fosfodiesterové vazby stejnou orientaci. Proto existuje polarita vlákna, rozlišující mezi 3 'a 5' koncem.
Podle konvence je struktura nukleových kyselin reprezentována 5 'koncem vlevo a 3' koncem vpravo.
Produkt RNA transkripce DNA je jednořetězcový pás, který se otáčí doprava, ve šroubovicové konformaci stohováním bází. Interakce mezi puriny je mnohem větší než interakce mezi dvěma pyrimidiny, vzhledem k jejich velikosti.
V RNA není možné mluvit o tradiční a referenční sekundární struktuře, jako je dvojitá šroubovice DNA. Trojrozměrná struktura každé molekuly RNA je jedinečná a složitá, srovnatelná se strukturou proteinů (logicky nemůžeme globalizovat strukturu proteinů).
Existují slabé interakce, které přispívají ke stabilizaci RNA, zejména stohování bází, kde jsou prstence umístěny jeden na druhém. Tento jev také přispívá ke stabilitě šroubovice DNA..
Pokud molekula RNA najde komplementární sekvenci, bude schopna se spojit a vytvořit dvouvláknovou strukturu, která se otočí doprava. Převládající forma je typu A; Pokud jde o formy Z, byly prokázány pouze v laboratoři, zatímco forma B nebyla pozorována.
Obecně existují krátké sekvence (jako je UUGG), které jsou umístěny na konci RNA a mají zvláštnost tvorby smyčky stabilní. Tato sekvence se účastní skládání trojrozměrné struktury RNA.
Kromě toho se mohou vodíkové vazby tvořit na jiných místech, než jsou typické párování bází (AU a CG). K jedné z těchto interakcí dochází mezi 2'-OH ribózy s jinými skupinami.
Elucidace různých struktur nalezených v RNA sloužila k prokázání mnoha funkcí této nukleové kyseliny..
Existují dvě třídy RNA: informační a funkční. První skupina zahrnuje RNA, které se účastní syntézy proteinů a fungují jako prostředníci v procesu; informační RNA jsou messengerové RNA.
Naproti tomu RNA patřící do druhé třídy, funkční, nevyvolává novou molekulu proteinu a samotná RNA je konečným produktem. Jedná se o transferové RNA a ribozomální RNA..
V savčích buňkách je 80% RNA ribozomální RNA, 15% přenosová RNA a pouze malá část odpovídá messengerové RNA. Tyto tři typy spolupracují na dosažení biosyntézy bílkovin..
Existují také malé jaderné RNA, malé cytoplazmatické RNA a mikroRNA. Každý z nejdůležitějších typů bude podrobně popsán níže:
U eukaryot je DNA omezena na jádro, zatímco syntéza proteinů probíhá v cytoplazmě buňky, kde se nacházejí ribozomy. Kvůli této prostorové separaci musí existovat mediátor, který přenáší zprávu z jádra do cytoplazmy a touto molekulou je poselská RNA..
Messenger RNA, zkráceně mRNA, je intermediární molekula, která obsahuje informace kódované v DNA a která specifikuje aminokyselinovou sekvenci, která povede k vzniku funkčního proteinu..
Termín messenger RNA navrhli v roce 1961 François Jacob a Jacques Monod k popisu části RNA, která přenášela zprávu z DNA na ribozomy..
Proces syntézy mRNA z řetězce DNA je známý jako transkripce a vyskytuje se rozdílně mezi prokaryoty a eukaryoty..
Genová exprese je řízena několika faktory a závisí na potřebách každé buňky. Přepis je rozdělen do tří fází: zahájení, prodloužení a ukončení..
Proces replikace DNA, ke kterému dochází v každém buněčném dělení, kopíruje celý chromozom. Proces transkripce je však mnohem selektivnější, zabývá se pouze zpracováním specifických segmentů řetězce DNA a nevyžaduje primer..
Na Escherichia coli -nejlépe studovaná bakterie v biologických vědách - transkripce začíná odvíjením dvojité šroubovice DNA a vzniká transkripční smyčka. Enzym RNA polymeráza je zodpovědný za syntézu RNA a jak transkripce pokračuje, řetězec DNA se vrací do původního tvaru.
Transkripce není zahájena na náhodných místech na molekule DNA; existují specializované stránky pro tento fenomén, zvané promotéry. Na E-coli RNA polymeráza je navázána několik párů bází nad cílovou oblastí.
Sekvence, kde jsou transkripční faktory spojené, jsou mezi různými druhy poměrně konzervativní. Jednou z nejznámějších promotorových sekvencí je TATA box.
Při prodloužení přidá enzym RNA polymeráza nové nukleotidy na 3'-OH konec ve směru 5 'až 3'. Hydroxylová skupina působí jako nukleofil a útočí na alfa fosfát přidávaného nukleotidu. Tato reakce uvolňuje pyrofosfát.
K syntéze messengerové RNA se používá pouze jeden z řetězců DNA, který se kopíruje ve směru 3 'až 5' (antiparalelní forma nového řetězce RNA). Nukleotid, který má být přidán, musí vyhovovat párování bází: páry U s A a G s C.
RNA polymeráza zastaví proces, když najde oblasti bohaté na cytosin a guanin. Nakonec je nová molekula messengerové RNA oddělena od komplexu.
U prokaryot může molekula messengerové RNA kódovat více než jeden protein.
Pokud mRNA kóduje výhradně protein nebo polypeptid, nazývá se to monocistronická mRNA, ale pokud kóduje více než jeden proteinový produkt, je mRNA polycistronická (všimněte si, že v této souvislosti termín cistron označuje gen).
U eukaryotických organismů je drtivá většina mRNA monocistronická a transkripční aparát je v této linii organismů mnohem složitější. Jsou charakterizovány tím, že mají tři RNA polymerázy, označené I, II a III, každá se specifickými funkcemi.
I má na starosti syntézu pre-rRNA, II syntetizuje messengerové RNA a některé speciální RNA. Nakonec je III zodpovědný za přenos RNA, ribozomální 5S a další malé RNA..
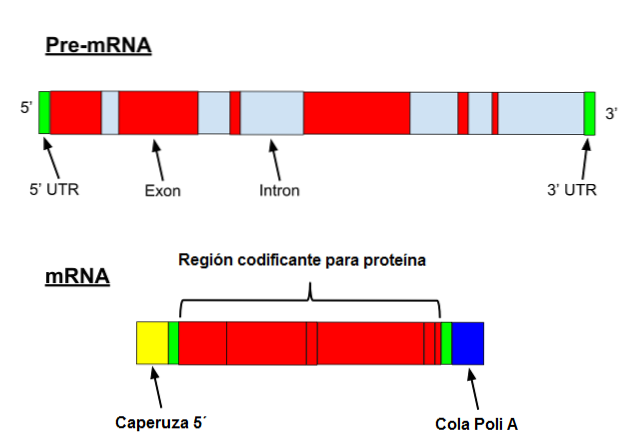
Messenger RNA prochází u eukaryotů řadou specifických modifikací. První zahrnuje přidání „čepičky“ na 5 'konec. Chemicky je uzávěrem 7-methylguanosinový zbytek připojený ke konci 5 ', 5'-trifosfátovou vazbou..
Funkcí této zóny je chránit RNA před možnou degradací ribonukleázami (enzymy, které štěpí RNA na menší složky).
Kromě toho dojde k odstranění 3 'konce a přidá se 80 až 250 adeninových zbytků. Tato struktura je známá jako polyA "ocas" a slouží jako vazebné místo pro různé proteiny. Když prokaryot získá polyA ocas, má tendenci stimulovat jeho degradaci..
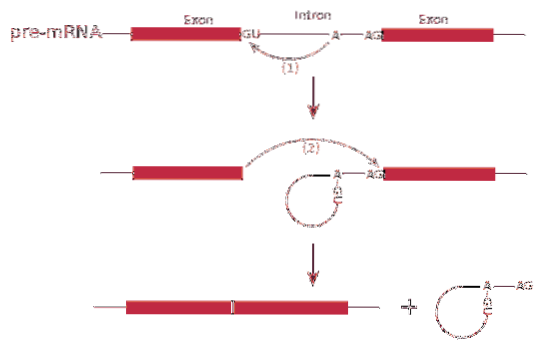
Na druhé straně je tento posel přepsán introny. Introny jsou sekvence DNA, které nejsou součástí genu, ale které „přerušují“ tuto sekvenci. Introny nejsou přeloženy, a proto musí být z posla odstraněny.
Většina genů obratlovců má introny, s výjimkou genů kódujících histony. Podobně se počet intronů v genu může pohybovat od několika do desítek..
Splicing Proces RNA nebo sestřihu zahrnuje odstranění intronů v messengerové RNA.
Některé introny nalezené v jaderných nebo mitochondriálních genech mohou provádět proces sestřih bez pomoci enzymů nebo ATP. Místo toho se proces provádí transesterifikačními reakcemi. Tento mechanismus byl objeven v řasinkách prvoků Tetrahymena thermophila.
Naproti tomu existuje další skupina poslů, kteří nejsou schopni zprostředkovat své vlastní sestřih, takže potřebují další strojní zařízení. Do této skupiny patří poměrně velké množství jaderných genů..
Proces sestřih je zprostředkován proteinovým komplexem nazývaným spliceosom nebo sestřihový komplex. Systém se skládá ze specializovaných komplexů RNA nazývaných malé nukleární ribonukleoproteiny (RNP)..
Existuje pět typů RNP: U1, U2, U4, U5 a U6, které se nacházejí v jádru a zprostředkovávají proces sestřih.
The sestřih může produkovat více než jeden typ bílkovin - toto je známé jako sestřih alternativní - protože exony jsou uspořádány odlišně a vytvářejí odrůdy poselských RNA.
Ribozomální RNA, zkráceně rRNA, se nachází v ribozomech a podílí se na biosyntéze proteinů. Proto je nezbytnou součástí všech buněk.
Ribozomální RNA se asociuje s molekulami proteinu (přibližně 100), čímž vznikají ribozomální předjednotky. Jsou klasifikovány v závislosti na jejich sedimentačním koeficientu, označeném písmenem S pro jednotky Svedberg..
Ribozom se skládá ze dvou částí: hlavní podjednotky a vedlejší podjednotky. Obě podjednotky se liší mezi prokaryoty a eukaryoty z hlediska koeficientu sedimentace..
Prokaryoty mají velkou podjednotku 50S a malou podjednotku 30S, zatímco u eukaryot je velká podjednotka 60S a malá 40S.
Geny, které kódují ribozomální RNA, jsou v jádru, což je zvláštní oblast jádra, která není ohraničena membránou. Ribozomální RNA jsou v této oblasti transkribovány RNA polymerázou I..
V buňkách, které syntetizují velké množství proteinů; jádro je prominentní struktura. Pokud však daná buňka nevyžaduje vysoký počet proteinových produktů, je jádro téměř nepostřehnutelnou strukturou.
60S velká ribozomální podjednotka je spojena s fragmenty 28S a 5,8S. Pokud jde o malou podjednotku (40S), je spojena s 18S.
U vyšších eukaryot je pre-rRNA kódována v transkripční jednotce 45S, která zahrnuje RNA polymerázu I. Tento transkript je zpracován na zralé ribosomální RNA 28S, 18S a 5,8S..
Jak syntéza pokračuje, pre-rRNA se asociuje s různými proteiny a tvoří částice ribonukleoproteinu. Prochází řadou následných modifikací, které zahrnují methylaci 2'-OH skupiny ribózy a přeměnu uridinových zbytků na pseudouridin..
Oblast, kde k těmto změnám dojde, je řízena více než 150 malými nukleolárními molekulami RNA, které mají schopnost se spojit s pre-rRNA..
Na rozdíl od zbytku pre-rRNA je 5S transkribován RNA polymerázou III v nukleoplazmě a ne uvnitř nukleolu. Poté, co je syntetizován, je přenesen do jádra, aby se sestavil s 28S a 5.8S, čímž se vytvoří ribozomální jednotky.
Na konci procesu montáže jsou podjednotky přeneseny do cytoplazmy přes jaderné póry..
Může se stát, že molekula messengerové RNA vytvoří několik proteinů současně, které se vážou na více než jeden ribozom. Jak postup překladu postupuje, konec posla se uvolní a může být vychytán jiným ribozomem a zahájit novou syntézu..
Z tohoto důvodu je běžné najít ribozomy seskupené (mezi 3 a 10) v jedné molekule messengerové RNA a tato skupina se nazývá polyribosom.
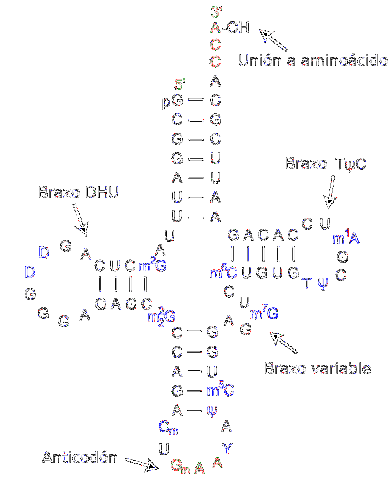
Transfer RNA je zodpovědná za přenos aminokyselin v průběhu procesu syntézy bílkovin. Jsou tvořeny přibližně 80 nukleotidy (ve srovnání s messengerovou RNA je to „malá“ molekula).
Struktura má záhyby a kříže připomínající trojramennou trojlístek. Na jednom z konců je umístěn adenylový kruh, kde hydroxylová skupina ribózy zprostředkovává spojení s transportovanou aminokyselinou.
Různé přenosové RNA se kombinují výhradně s jednou z dvaceti aminokyselin, které tvoří proteiny; jinými slovy, je to nosič, který nese stavební kameny bílkovin. Komplex přenosu RNA společně s aminokyselinou se nazývá aminoacyl-tRNA.
Kromě toho v procesu translace - ke kterému dochází díky ribozomům - každá přenosová RNA rozpoznává specifický kodon v messengerové RNA. Po rozpoznání se uvolní odpovídající aminokyselina a stane se součástí syntetizovaného peptidu.
Aby rozpoznala typ aminokyseliny, která musí být dodána, má RNA „antikodon“ umístěný ve střední oblasti molekuly. Tento antikodon je schopen tvořit vodíkové vazby s komplementárními bázemi přítomnými v messengerové DNA..
MicroRNA nebo miRNA jsou typem krátké, jednořetězcové RNA, mezi 21 a 23 nukleotidy, jejichž funkcí je regulovat expresi genů. Vzhledem k tomu, že se nepřekládá na protein, často se nazývá nekódující RNA..
Stejně jako jiné typy RNA je zpracování mikroRNA složité a zahrnuje řadu proteinů.
MikroRNA vznikají z delších prekurzorů nazývaných mi-pri-RNA, odvozených z prvního transkriptu genu. V jádru buňky jsou tyto prekurzory modifikovány v mikroprocesorovém komplexu a výsledkem je pre-miRNA..
Pre-miRNA jsou 70 nukleotidové vlásenky, které pokračují ve zpracování v cytoplazmě pomocí enzymu zvaného Dicer, který sestavuje komplex umlčování vyvolaný RNA (RISC) a nakonec je syntetizována miRNA..
Tyto RNA jsou schopné regulovat expresi genů, protože jsou komplementární se specifickými messengerovými RNA. Spojením se svým cílem jsou miRNA schopné potlačit posla nebo ho dokonce degradovat. V důsledku toho nemůže ribozom překládat uvedený přepis..
Jedním konkrétním typem mikroRNA jsou malé interferující RNA (siRNA), nazývané také umlčující RNA. Jsou to krátké RNA mezi 20 až 25 nukleotidy, které brání expresi určitých genů.
Jsou velmi slibnými nástroji pro výzkum, protože umožňují umlčet požadovaný gen a studovat jeho možnou funkci..
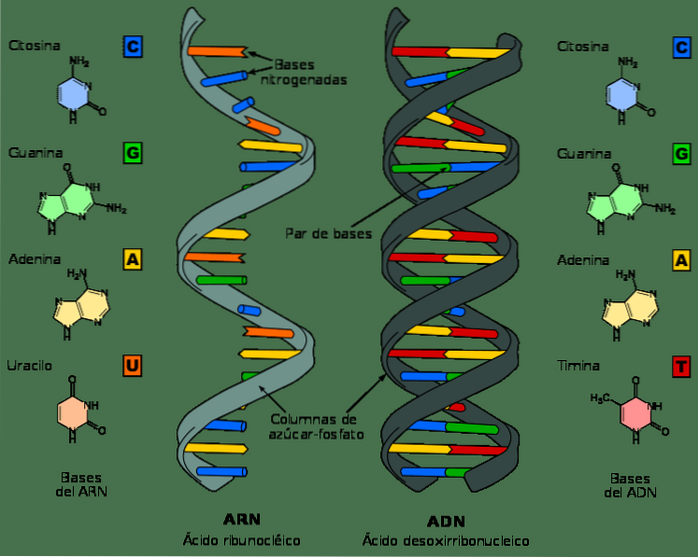
Ačkoli DNA a RNA jsou nukleové kyseliny a na první pohled mohou vypadat velmi podobně, liší se v několika svých chemických a strukturních vlastnostech. DNA je dvojpásmová molekula, zatímco RNA je jednopásmová.
Výsledkem je, že RNA je univerzálnější molekula a může nabývat široké škály trojrozměrných tvarů. Některé viry však mají ve svém genetickém materiálu dvoupásmovou RNA..
V RNA nukleotidech je molekulou cukru ribóza, zatímco v DNA deoxyribóza, která se liší pouze v přítomnosti atomu kyslíku.
Fosfodiesterová vazba v páteři DNA a RNA je náchylná k pomalému procesu hydrolýzy bez přítomnosti enzymů. Za alkalických podmínek RNA rychle hydrolyzuje - díky extra hydroxylové skupině - zatímco DNA ne.
Podobně dusíkatými bázemi, které tvoří nukleotidy v DNA, jsou guanin, adenin, thymin a cytosin; na rozdíl od toho je v RNA thymin nahrazen uracilem. Uracil se může spárovat s adeninem, stejně jako thymin v DNA.
RNA je jediná známá molekula schopná uchovávat informace a současně katalyzovat chemické reakce; Z tohoto důvodu několik autorů navrhuje, aby molekula RNA byla rozhodující pro vznik života. Překvapivě jsou substráty ribozomu další molekuly RNA.
Objev ribozymů vedl k biochemické redefinici „enzymu“ - protože dříve byl tento termín používán výhradně pro proteiny s katalytickou aktivitou - a pomohl podpořit scénář, kdy první formy života používaly jako genetický materiál pouze RNA.


Zatím žádné komentáře