The erytropoéza Je to proces, při kterém se tvoří červené krvinky nebo erytrocyty. Tyto krevní buňky mají u lidí průměrnou životnost 4 měsíce a nejsou schopné samy se reprodukovat. Z tohoto důvodu musí být vytvořeny nové erytrocyty, které nahradí ty, které zemřou nebo se ztratí při krvácení..
U mužů je počet červených krvinek přibližně 54 milionů na mililiter, zatímco u žen je to o něco méně (48 milionů). Asi 10 milionů erytrocytů je ztraceno denně, takže je třeba vyměnit podobné množství.
Erytrocyty se tvoří z nukleovaných erytroblastů přítomných v červené kostní dřeni savců, zatímco u jiných obratlovců se produkují hlavně v ledvinách a slezině.
Když dosáhnou konce svých dnů, rozdělí se; pak je obklopují buňky zvané makrofágy. Tyto makrofágy jsou přítomny v játrech, červené kostní dřeni a slezině..
Když jsou červené krvinky zničeny, železo se recykluje a znovu se použije, zatímco zbytek hemoglobinu se přemění na žlučový pigment zvaný bilirubin..
Erytropoéza je stimulována hormonem nazývaným erytropoetin, ale proces je regulován různými faktory, jako je například teplota, tlak kyslíku..
Rejstřík článků
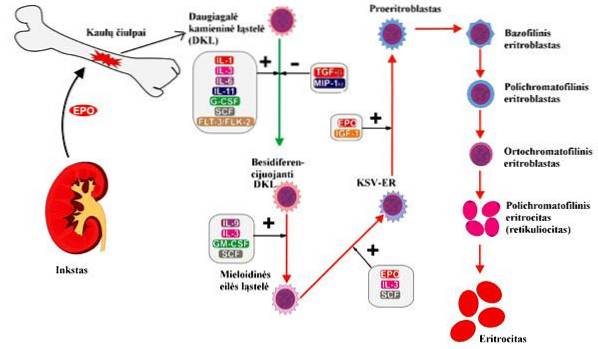
U dospělých organismů se erytropoéza vyskytuje na specializovaných místech v červené kostní dřeni nazývaných erytroblastické ostrovy. K tvorbě erytrocytů musí dojít k několika procesům, od buněčné proliferace po zrání červených krvinek, které procházejí různými stadii buněčné diferenciace..
Jak buňky procházejí mitotickými děleními, zmenšuje se jejich velikost a velikost jejich jádra, stejně jako kondenzace chromatinu a hemoglobinizace. Navíc se vzdalují od oblasti původu.
V závěrečných fázích ztratí jádro a další organely a vstoupí do oběhu migrujícími přes cytoplazmatické póry endotelových buněk..
Někteří autoři rozdělují celý proces erytropoézy do dvou fází, první buněčné proliferace a diferenciace; zatímco jiní dělí proces na základě specifických charakteristik buňky v každé fázi, když jsou pozorováni pomocí Wrightova skvrn. Na základě toho jsou stádia erytropoézy:
Jsou to první buňky citlivé na erytropoetin, někteří autoři je pro svou zkratku v angličtině nazývají myeloidní progenitory, nebo také BFU-E. Jsou charakterizovány expresí povrchových antigenů, jako je CD34, a také přítomností erytropoetinových receptorů v malých množstvích.
Zkráceny v angličtině jako CFU-E, jsou schopné produkovat malé kolonie erytroblastů. Další charakteristikou těchto buněk je, že množství erytropoetinových receptorů je mnohem vyšší než v jednotkách tvořících kolonie..
Považováno za první fázi zrání erytrocytů. Vyznačují se velkou velikostí (14 až 19 µm podle některých autorů, až 25 µm podle jiných). Jádro je zaoblené a také obsahuje nukleoly a bohatý chromatin.
Považováno za první fázi zrání erytrocytů. Vyznačují se velkou velikostí (14 až 19 µm podle některých autorů, až 25 µm podle jiných). Jádro je velké, zaoblené, s chromatinem uspořádaným ve formě vláken a 2 nebo 3 nukleolů.
V této fázi začíná absorpce železa v plazmě. Mají poločas 20 hodin, aby ustoupili mitózou do další fáze.
Také se jim říká normoblasty, jsou menší než jejich předchůdci. Tyto buňky se barví modře vitálním zabarvením, to znamená, že jsou bazofilní. Jádro je kondenzované, nukleoly zmizely a mají velké množství ribozomů. V této fázi začíná syntéza hemoglobinu.
Na začátku jsou známé jako bazofilní erytroblasty typu I a po mitotickém dělení se transformují na typ II, které zůstávají bazofily a vykazují vyšší syntézu hemoglobinu. Přibližná doba trvání obou buněk je podobná jako u proerythroblastů.
Jsou tvořeny mitotickým dělením bazofilních erytroblastů typu II a jsou posledními buňkami schopnými dělení mitózou. Jejich velikost se pohybuje od 8 do 12 µm a mají zaoblené a kondenzované jádro.
Cytoplazma těchto buněčných skvrn vede šedě s Wrightovou barvou. Má vysokou koncentraci hemoglobinu a počet ribozomů zůstává vysoký.
6-ortochromatické erytroblasty
Barva těchto buněk je růžová nebo červená kvůli množství hemoglobinu, které mají. Jeho velikost je o něco menší než u jeho prekurzorů (7 až 10 µm) a má malé jádro, které bude vyloučeno exocytózou, až buňky dozrají..
Vznikají diferenciací ortochromních erytroblastů, ztrácejí organely a plní svou cytoplazmu hemoglobinem. Zůstávají v červené kostní dřeni dva až tři dny, dokud migrují do krve, kde dokončí své zrání..
Jsou to zralé vytvořené prvky, konečný produkt erytropoézy a které se tvoří zráním retikulocytů. Mají bikonkávní tvar kvůli absenci jádra a interakci mezi cytoskeletem erytrocytů a dvěma proteiny zvanými spektrin a aktin.
Jsou to nejhojnější krvinky, jsou tvořeny retikulocyty. U savců mají bikonkávní tvar kvůli absenci jádra a interakci mezi cytoskeletem erytrocytů a dvěma proteiny zvanými spektrin a aktin. U ostatních obratlovců jsou zaoblené a zachovávají si jádro.
Přestože erytropoetin stimuluje tvorbu červených krvinek ke zlepšení schopnosti krve přenášet kyslík, existuje několik základních mechanismů pro regulaci této tvorby, včetně:
Koncentrace kyslíku v krvi reguluje erytropoézu. Když je tato koncentrace v průtoku krve ledvinami velmi nízká, stimuluje se produkce červených krvinek..
Tato nízká koncentrace O2 v tkáni může nastat v důsledku hypoxémie, anémie, ischemie ledvin nebo když je afinita hemoglobinu k tomuto plynu vyšší než obvykle..
Miescher v roce 1893 jako první navrhl vztah mezi hypoxií tkáně a erytropoézou. Tato hypoxie však přímo nestimuluje kostní dřeň k produkci červených krvinek, jak navrhl Miescher. Naopak, indukuje ledviny k produkci hormonu erytropoetinu.
Produkce erytropoetinu v důsledku tkáňové hypoxie je geneticky regulovaná a receptory, které tuto hypoxii detekují, se nacházejí v ledvinách. Produkce erytropoetinu se také zvyšuje v důsledku poklesu parciálního tlaku kyslíku ve tkáni po krvácení..
Buňky, které produkují erytropoetin, se nacházejí v ledvinách a játrech. Zvýšení produkce tohoto hormonu během anémie je způsobeno zvýšením počtu buněk, které jej produkují..
Testosteron nepřímo reguluje erytropoézu regulací hladin železa v krvi. Tento hormon působí přímo na působení cytoplazmatického proteinu zvaného BMP-Smad (kostní morfogenetický protein-Smad) v hepatocytech.
Díky působení testosteronu je transkripce hepcidinu potlačena. Tento hepcidin zabraňuje průchodu železa z buněk do plazmy z makrofágů, které recyklují železo, což vede k drastickému poklesu železa v krvi.
Když dojde k hypoferremii, dojde k inhibici erytropoetinu, protože nebude existovat železo pro produkci erytrocytů.
Bylo prokázáno, že teplota má vliv na erytripoézu. Vystavení velmi nízkým teplotám způsobuje potřebu vytvářet teplo v tkaninách.
To vyžaduje zvýšení počtu erytrocytů za účelem dodávky kyslíku do periferních tkání. Není však zcela jasné, jak k tomuto typu regulace dochází..
Zdá se, že neurony centrálního nervového systému produkují erytropoetin, aby se chránily před ischemickým poškozením a apoptózou. Vědci to však zatím nedokázali ověřit..
Činidla stimulující erytropoézu (ESA) jsou látky odpovědné za stimulaci produkce erytrocytů. Erytropoetin je hormon přirozeně odpovědný za tento proces, ale existují i syntetické produkty s podobnými vlastnostmi.
Erytropoetin je hormon syntetizovaný hlavně v ledvinách. V raných fázích vývoje se játra také podílejí na aktivní produkci erytropoetinu. Jak však pokračuje vývoj, druhý orgán se do procesu zapojuje méně..
Erytrocyt začíná dispergovat receptory erytropoetinu na povrchu membrány. Erytropoetin aktivuje řadu mezibuněčných signálních transdukčních kaskád, které zpočátku produkují syntézu hemoglobinu a způsobují, že retikulocyty působí rychleji a jsou uvolňovány do oběhu..
Umělé ESA se dělí na generace (první až třetí) v závislosti na datu jejich vytvoření a uvedení na trh. Strukturálně a funkčně podobné erytropoetinu.
ESA první generace jsou známé jako epoetin alfa, beta a delta. První dva jsou produkovány rekombinací ze zvířecích buněk a mají poločas asi 8 hodin v těle. Epoetinová delta se mezitím syntetizuje z lidských buněk.
Darbepoetin alfa je druhá generace ESA, vyráběná z buněk čínských křečků pomocí technologie zvané rekombinantní DNA. Má poločas rozpadu více než třikrát delší než u ESA první generace. Stejně jako u epoetinů někteří vysoce výkonní sportovci užívali darbepoetin jako dopingový prostředek.
Continuous Erythropoetin Receptor Activator, nebo CERA pro jeho zkratku v angličtině, je obecný název pro třetí generace ESA. Nesnaží se simulovat strukturu a funkci erytropoetinu, ale spíše stimulují receptor pro erytropoetin, čímž zvyšují jeho účinky..
Jeho poločas je několik týdnů místo hodin, stejně jako u předchozích léků. Komerčně se používá od roku 2008, jeho nezákonné použití při sportovních aktivitách však zřejmě pochází dva nebo tři roky před jeho legální komercializací.
K neúčinné nebo neúčinné erytropoéze dochází, když jsou vytvořené červené krvinky poškozené a obvykle jsou zničeny před nebo krátce po opuštění kostní dřeně..
Neúčinná erytropoéza může být způsobena vadami v syntéze nukleových kyselin, hemové skupiny nebo globinů. Tyto vady způsobují různé typy anémie.
V tomto případě je nedostatek kyseliny listové a kobalaminu, syntéza DNA je inhibována v jádru buněk promotoru erytrocytů, takže se nemohou mitoticky dělit. Cytoplazma sama o sobě zvětšuje svůj objem (makrocytóza) a vytváří velkou buňku zvanou megaloblast.
V těchto případech vzniká řada anémií zvaných megaloblastické anémie, z nichž nejčastější je perniciózní anémie. U tohoto onemocnění nedochází k absorpci vitaminu B12 v tenkém střevě.
Mezi další příčiny megaloblastické anémie patří nemoci zažívacího traktu, malabsorpce, nedostatek kyseliny listové a některé léky.
Mezi příznaky tohoto typu anémie patří abnormální bledost, podrážděnost, ztráta chuti k jídlu, průjem, potíže s chůzí nebo svalová slabost. V závislosti na příčině může být léčen doplňky vitaminu nebo kyseliny listové..
Neúčinná erytropoéza způsobená nedostatkem syntézy železa může způsobit dva typy anémie; mikrocytární anémie s nedostatkem železa a sideroblastická anémie.
Mikrocytární anémie je známá jako skupina anemií charakterizovaná malými a světle červenými krvinkami, mohou mít různý původ, včetně talasémie a neúčinné erytropoézy.
Hladina železa a hemosiderinu je u sideroblastické anémie velmi vysoká. Hemosiderin je žlutý pigment odvozený od hemoglobinu a objevuje se, když jsou hladiny kovu vyšší než obvykle. Tento typ anémie způsobuje smrt bazofilů v červené kostní dřeni a nedochází k syntéze hemoglobinu.
Říká se tomu sideroblastická anémie, protože erytrocyty se vyvíjejí abnormálně v důsledku hromadění železa ve formě granulí, které dostává název sideroblastů. Sideroblastická anémie může být vrozená nebo může být sekundární a může mít různé příčiny.
V tomto případě se objeví srpkovitá anémie a beta talasémie. Srpkovitá anémie je také známá jako srpkovitá anémie. Je produkován genetickou mutací, která vede k substituci kyseliny glutamové valinem během syntézy beta globinu.
Díky této substituci klesá afinita hemoglobinu ke kyslíku a dochází k atrofii erytrocytů, která místo normálního bikonkávního tvaru disku získává srpkovitý tvar. Pacient se srpkovitou anémií je náchylný k mikroinfarktům a hemolýze.
Thalasémie je onemocnění způsobené nedostatečným genetickým kódováním α- a β-globinů, které vede k předčasné smrti erytrocytů. Existuje asi sto různých mutací, které mohou způsobit talasemii s různým stupněm závažnosti.



Zatím žádné komentáře