The fosfatidylethanolamin (PE) je glycerofosfolipid hojně se vyskytující v plazmatických membránách prokaryotických organismů. Naopak v eukaryotických buněčných membránách se jedná o druhý nejhojnější glycerofosfolipid na vnitřní straně plazmatické membrány po fosfatidylcholinu..
Navzdory nadbytku fosfatidylethanolaminu jeho nadbytek závisí nejen na typu buňky, ale také na kompartmentu a okamžiku konkrétního uvažovaného životního cyklu buňky..
Biologické membrány jsou bariéry, které definují buněčné organismy. Nejen, že mají ochranné a izolační funkce, ale jsou také klíčem k vytvoření proteinů, které pro své optimální fungování vyžadují hydrofobní prostředí..
Eukaryota i prokaryota mají membrány složené převážně z glycerofosfolipidů a v menší míře sfingolipidů a sterolů..
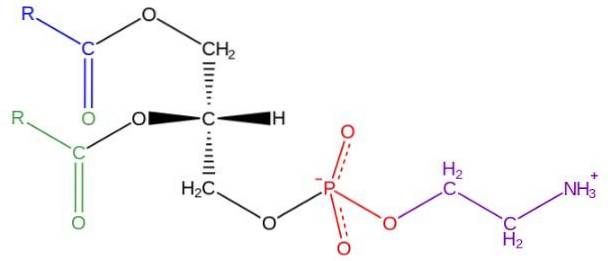
Glycerofosfolipidy jsou amfipatické molekuly strukturované na základním řetězci L-glycerolu, který je esterifikován v polohách sn-1 a sn-2 dvěma mastnými kyselinami s různou délkou a stupněm nasycení. V hydroxylu polohy sn-3 je esterifikován fosfátovou skupinou, ke které lze následně připojit různé typy molekul, které vedou k různým třídám glycerofosfolipidů.
V buněčném světě existuje široká škála glycerofosfolipidů, ale nejhojnější jsou fosfatidylcholin (PC), fosfatidylethanolamin (PE), fosfatidylserin (PS), fosfatidylinositol (PI), kyselina fosfatidová (PA), fosfatidylglycerol (PG) a kardiolipin. (CL).
Rejstřík článků
Strukturu fosfatidylethanolaminu objevil Baer a kol. V roce 1952. Jak bylo experimentálně stanoveno pro všechny glycerofosfolipidy, fosfatidylethanolamin je tvořen glycerolovou molekulou esterifikovanou v polohách sn-1 a sn-2 s kyselými řetězci mastnými s 16 až 20 atomy uhlíku.
Mastné kyseliny esterifikované v hydroxylové skupině sn-1 jsou obecně nasycené (bez dvojných vazeb) s maximální délkou 18 atomů uhlíku, zatímco řetězce spojené v poloze sn-2 jsou delší a s jednou nebo více nenasycenostmi (dvojné vazby).
Stupeň nasycení těchto řetězců přispívá k pružnosti membrány, což má velký vliv na inzerci a sekvestraci proteinů do dvojvrstvy..
Fosfatidylethanolamin je považován za nelamelární glycerofosfolipid, protože má kuželovitý geometrický tvar. Tento tvar je dán malou velikostí jeho polární skupiny nebo „hlavy“ ve vztahu k velikosti řetězců mastných kyselin, které tvoří hydrofobní „ocasy“.
„Hlava“ nebo polární skupina fosfatidylethanolaminu má zwitteriontový charakter, to znamená, že má skupiny, které mohou být za určitých podmínek pH pozitivně a negativně nabité..
Tato vlastnost mu umožňuje vodíkovou vazbu s velkým počtem aminokyselinových zbytků a jeho distribuce náboje je podstatným determinantem pro topologii domény mnoha integrálních membránových proteinů..
V eukaryotických buňkách je syntéza strukturních lipidů geograficky omezená, hlavním biosyntetickým místem je endoplazmatické retikulum (ER) a v menší míře Golgiho aparát..
Existují čtyři nezávislé biosyntetické dráhy pro výrobu fosfatidylethanolaminů: (1) cesta CDP-ethanolamin, známá také jako Kennedyho dráha; (2) dráha PSD pro dekarboxylaci fosfatidylserinu (PS); (3) acylace lyso-PE a (4) reakce na změnu báze polární skupiny jiných glycerofosfolipidů.
Biosyntéza fosfatidylethanolaminu touto cestou je omezena na ER a bylo prokázáno, že v jaterních buňkách křečka je hlavní cestou produkce. Skládá se ze tří po sobě jdoucích enzymatických kroků katalyzovaných třemi různými enzymy.
V prvním kroku se fosfoethanolamin a ADP vyrábějí díky působení ethanolamin kinázy, která katalyzuje na ATP závislou fosforylaci ethanolaminu..
Na rozdíl od rostlin nejsou savci ani kvasinky schopni tento substrát produkovat, proto musí být konzumován ve stravě nebo získáván degradací již existujících molekul fosfatidylethanolaminů nebo sfingosinů..
Fosfoethanolamin používá CTP: fosfoethanolamin cytidyltransferáza (ET) k vytvoření vysokoenergetické sloučeniny CDP: ethanolamin a anorganický fosfát.
1,2-Diacylglycerol ethanolamin fosfotransferáza (ETP) využívá energii obsaženou v CDP-ethanolaminové vazbě k kovalentní vazbě ethanolaminu na molekulu diacylglycerolu s vloženou membránou, čímž vzniká fosfatidylethanolamin..
Tato cesta funguje jak u prokaryot, tak u kvasinek a savců. U bakterií se vyskytuje v plazmatické membráně, ale u eukaryot se vyskytuje v oblasti endoplazmatického retikula, která úzce souvisí s mitochondriální membránou.
U savců je dráha katalyzována jediným enzymem, fosfatidylserin dekarboxylázou (PSD1p), který je zality v mitochondriální membráně, jejíž gen je kódován jádrem. Reakce zahrnuje dekarboxylaci PS na fosfatidylethanolamin.
Zbývající dvě cesty (acylace PE-lyso a výměna vápníku závislá na polárních skupinách) se vyskytují v endoplazmatickém retikulu, ale významně nepřispívají k celkové produkci fosfatidylethanolaminů v eukaryotických buňkách..
Glycerofosfolipidy mají v buňce tři hlavní funkce, mezi kterými vynikají strukturální funkce, skladování energie a buněčná signalizace..
Fosfatidylethanolamin je spojen s ukotvením, stabilizací a složením více membránových proteinů, stejně jako s konformačními změnami nezbytnými pro fungování mnoha enzymů.
Existují experimentální důkazy, které navrhují fosfatidylethanolamin jako rozhodující glycerofosfolipid v pozdním stadiu telofázy, během tvorby kontraktilního kruhu a vzniku fragmoplastu, který umožňuje dělení membrány dvou dceřiných buněk..
Má také důležitou roli ve všech procesech fúze a štěpení (spojení a separace) membrán endoplazmatického retikula i Golgiho aparátu..
U E. coli se prokázalo, že fosfatidylethanolamin je nezbytný pro správné skládání a funkci enzymu laktóza permeáza, a proto se předpokládá, že hraje roli jako molekulární „chaperon“.
Fosfatidylethanolamin je hlavním dárcem molekuly ethanolaminu nezbytnou pro posttranslační modifikaci mnoha proteinů, jako jsou kotvy GPI.
Tento glycerofosfolipid je předchůdcem mnoha molekul s enzymatickou aktivitou. Kromě toho mohou molekuly odvozené z jeho metabolismu, stejně jako diacylglycerol, kyselina fosfatidová a některé mastné kyseliny, působit jako druzí poslové. Kromě toho je důležitým substrátem pro výrobu fosfatidylcholinu..



Zatím žádné komentáře