The globosidy Jsou typem sfingolipidů patřících do heterogenní rodiny glykosfingolipidů a vyznačují se tím, že mají ve svých strukturách polární skupinu složenou ze složitých strukturovaných glykanů spojených s ceramidovou kostrou B-glykosidovou vazbou.
Jsou klasifikovány v rámci „glóbusové“ řady glykosfingolipidů přítomností centrální struktury obecné formy Galα4Galβ4GlcβCer a jejich nomenklatura je obecně založena na počtu a typu sladkých zbytků polárních hlav.
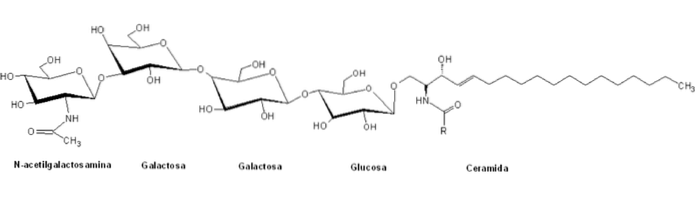
Na rozdíl od jiných sfingolipidů jsou globosidy normální složkou buněčných membrán nervových systémových orgánů mnoha savců. Například ledviny, střeva, plíce, nadledviny a červené krvinky.
Stejně jako všechny membránové lipidy mají globosidy důležité strukturální funkce při tvorbě a řazení lipidových dvojvrstev..
Avšak na rozdíl od jejich kyselých nebo fosforylovaných protějšků funkce globosidů nesouvisí tolik s produkcí signálních molekul, ale spíše s jejich účastí jako součásti glykokonjugátů v plazmatické membráně..
Rejstřík článků
Sdílejí některé strukturální a funkční podobnosti s ostatními členy skupiny glukosfingolipidů: cerebrosidy, gangliosidy a sulfatidy; včetně složení hlavní kostry a vedlejších produktů jejího metabolismu.
Globosidy se však liší od kyselých glykosfingolipidů (jako jsou gangliosidy), pokud jde o náboj jejich sacharidových polárních skupin, protože jsou elektricky neutrální při fyziologickém pH, což se zdá mít silné důsledky pro jejich funkce jako součásti extracelulární matrice..
Tyto skupiny polárních hlav mají obvykle více než dvě molekuly cukru, mezi nimiž jsou obvykle D-glukóza, D-galaktóza a N-acetyl-D-galaktosamin, a v menší míře fukóza a N-acetylglukosamin.
Stejně jako u jiných sfingolipidů mohou být globosidy velmi rozmanité molekuly, a to buď s přihlédnutím k více kombinacím mastných kyselin připojených ke kostře sfingosinů, nebo k možným variacím oligosacharidových řetězců hydrofilní části..
Cesta začíná syntézou ceramidu v endoplazmatickém retikulu (ER). Kostra sfingosinu se nejprve vytvoří kondenzací L-serinu a palmitoyl-CoA.
Ceramid je následně generován působením enzymů ceramid syntázy, které kondenzují další molekulu mastné kyseliny-CoA s páteří sfingosinu na uhlíku v poloze 2.
Stále v ER mohou být vyrobené ceramidy modifikovány přidáním zbytku galaktózy za vzniku galakto ceramidů (GalCer), nebo mohou být místo toho transportovány do Golgiho komplexu buď působením ceramidových přenosových proteinů (CERT) nebo pomocí vezikulárního transportu.
V komplexu Golgi lze ceramidy glykosylovat za vzniku gluko ceramidů (GlcCer).
GlcCer se vyrábí na cytosolické tváři raných Golgi. Poté může být transportován do luminální strany komplexu a následně může být glykosylován specifickými glykosidázovými enzymy, které generují složitější glykosfingolipidy..
Společné prekurzory všech glykosfingolipidů jsou syntetizovány v komplexu Golgi působením glykosyltransferáz z GalCer nebo GlcCer.
Tyto enzymy přenášejí specifické sacharidy z příslušných nukleotidových cukrů: UDP-glukóza, UDP-galaktóza, CMP-kyselina sialová atd..
Když GlcCer prochází vezikulárním transportním systémem Golgiho, je galaktosylován za vzniku laktosylceramidu (LacCer). LacCer je větev, ze které se syntetizují prekurzory ostatních glykosfingolipidů, tj. Molekula, do které se následně přidávají neutrálnější polární zbytky cukru. Tyto reakce jsou katalyzovány specifickými globosid syntázami.
Tyto lipidy se nacházejí hlavně v lidských tkáních. Stejně jako mnoho glykosfingolipidů jsou globosidy obohaceny na vnější straně plazmatické membrány mnoha buněk..
Jsou zvláště důležité v lidských erytrocytech, kde představují hlavní typ glykolipidů na povrchu buněk..
Kromě toho, jak je uvedeno výše, jsou součástí souboru glykokonjugátů plazmatických membrán mnoha nervových orgánů, zejména ledvin..
Funkce globosidů dosud nebyly zcela objasněny, ale je známo, že některé druhy zvyšují buněčnou proliferaci a motilitu, na rozdíl od inhibice těchto událostí způsobené některými gangliosidy..
Tetra-glykosylovaný globosid, Gb4 (GalNAcβ3Galα4Galβ4GlcβCer), funguje při místně citlivém rozpoznávání strukturálních poruch erytrocytů během procesu buněčné adheze..
Nedávné studie určily účast Gb4 na aktivaci ERK proteinů v buněčných liniích karcinomu, což by mohlo znamenat jeho účast na zahájení nádoru. Tyto proteiny patří do signalizační kaskády mitogenem aktivované proteinkinázy (MAPK), skládající se z prvků Raf, MEK a ERK..
Byla popsána jejich účast jako receptorů pro některé bakteriální toxiny rodiny Shiga, konkrétně globosid Gb3 (Galα4Galβ4GlcβCer), také známý jako CD77, exprimovaný v nezralých B buňkách; také jako receptory pro faktor adheze na HIV (gp120) a zdá se, že mají důsledky pro určité typy rakoviny a jiných nemocí.
U lidí existuje mnoho typů lipidózy. Globosidy a jejich metabolické dráhy souvisejí zejména se dvěma chorobami: Fabryho chorobou a Sandhoffovou chorobou..
Vztahuje se na zděděnou systémovou poruchu spojenou s pohlavím, poprvé pozorovanou u pacientů s více fialovými skvrnami v pupeční oblasti. Ovlivňuje orgány, jako jsou ledviny, srdce, oči, končetiny, část gastrointestinálního a nervového systému.
Je to produkt metabolického defektu v enzymu ceramid trihexosidáza, který je zodpovědný za hydrolýzu trihexosiceramidu, meziproduktu v katabolismu globosidů a gangliosidů, což způsobuje akumulaci těchto glykolipidů ve tkáních.
Tato patologie byla původně popsána jako varianta Tay-Sachsovy choroby související s metabolizmem gangliosidů, ale také to představuje akumulaci globosidů ve vnitřnostech. Jedná se o dědičnou poruchu s autozomálně recesivními vzory, která postupně ničí neurony a míchu.
Souvisí to s absencí forem A a B enzymu β-N-acetylhexosaminidáza v důsledku mutací v genu HEXB. Tyto enzymy jsou zodpovědné za jeden z degradačních kroků některých glykosfingolipidů.



Zatím žádné komentáře