The nukleosom je základní jednotkou balení DNA v eukaryotických organismech. Jedná se tedy o nejmenší prvek pro kompresi chromatinu.
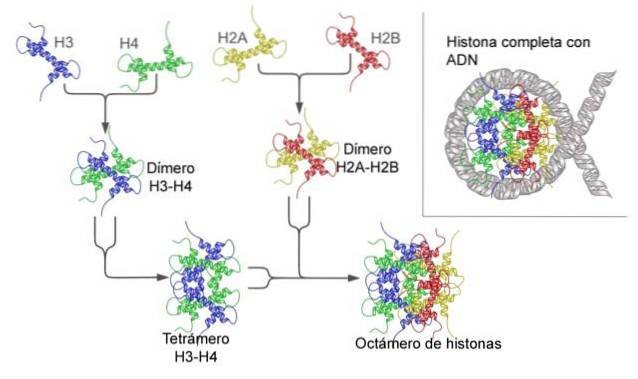
Nukleosom je postaven jako oktamer proteinů nazývaných histony, nebo struktura ve tvaru bubnu, na které je navinuto asi 140 nt DNA, což dělá téměř dvě kompletní otočení..
Navíc je za část nukleosomu považováno dalších 40-80 nt DNA a je to frakce DNA, která umožňuje fyzickou kontinuitu mezi jedním nukleosomem a jiným ve složitějších strukturách chromatinu (jako je 30nm vlákno chromatinu)..
Histonový kód byl jedním z prvních molekulárně pochopených epigenetických řídicích prvků..
Rejstřík článků
Nukleosomy umožňují:
V nejzákladnějším smyslu jsou nukleosomy tvořeny DNA a bílkovinami. DNA může být prakticky jakákoli dvoupásmová DNA přítomná v jádru eukaryotické buňky, zatímco nukleosomální proteiny všechny patří do sady proteinů nazývaných histony..
Histony jsou malé proteiny s vysokým obsahem bazických aminokyselinových zbytků; To umožňuje působit proti vysokému negativnímu náboji DNA a navázat účinnou fyzickou interakci mezi dvěma molekulami bez dosažení tuhosti kovalentní chemické vazby..
Histony tvoří bubenovitý oktamer se dvěma kopiemi nebo monomery každého z histonů H2A, H2B, H3 a H4. DNA provede téměř dvě úplná otočení po stranách oktameru a poté pokračuje s frakcí linkerové DNA, která se asociuje s histonem H1, aby se vrátila a poskytla dvě úplné otočení na dalším histonovém oktameru..
Sada oktamerů, asociovaná DNA a její odpovídající linkerová DNA, je nukleosom.
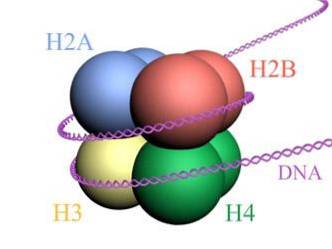
Genomická DNA je tvořena extrémně dlouhými molekulami (více než jeden metr v případě člověka, s ohledem na všechny jejich chromozomy), které musí být zhutněny a uspořádány v extrémně malém jádru.
První krok v tomto zhutnění se provádí tvorbou nukleosomů. Pouze s tímto krokem je DNA zhutněna asi 75krát.
To vede k lineárnímu vláknu, ze kterého jsou sestaveny následné úrovně zhutnění chromatinu: 30 nm vlákno, smyčky a smyčky smyček..
Když se buňka dělí buď mitózou nebo meiózou, konečným stupněm zhutnění je samotný mitotický nebo meiotický chromozom..
Skutečnost, že histonové oktamery a DNA interagují elektrostaticky, částečně vysvětluje jejich efektivní asociaci, aniž by došlo ke ztrátě tekutosti potřebné k tomu, aby se nukleosomy staly dynamickými prvky pro zhutňování a dekomprimaci chromatinu..
Existuje však ještě překvapivější prvek interakce: N-koncové konce histonů jsou vystaveny mimo vnitřek oktameru, který je kompaktnější a inertnější..
Tyto konce nejen fyzicky interagují s DNA, ale také procházejí řadou kovalentních modifikací, na kterých bude záviset stupeň zhutnění chromatinu a exprese asociované DNA..
Sada kovalentních modifikací, pokud jde o typ a počet, je mimo jiné kolektivně známá jako histonový kód. Tyto modifikace zahrnují fosforylaci, methylaci, acetylaci, ubikvitinaci a sumoylaci argininových a lysinových zbytků na N-koncích histonů..
Každá změna, společně s dalšími v rámci stejné molekuly nebo ve zbytcích jiných histonů, zejména histonů H3, určí nebo nezohlední expresi asociované DNA, jakož i stupeň zhutnění chromatinu..
Obecně se například ukázalo, že hypermethylované a hypoacetylované histony určují, že asociovaná DNA není exprimována a že chromatin je přítomen v kompaktnějším stavu (heterochromatický, a tedy neaktivní)..
Naproti tomu euchromatická DNA (méně kompaktní a geneticky aktivní) je spojena s chromatinem, jehož histony jsou hyperacetylované a hypomethylované..
Již jsme viděli, že stav kovalentní modifikace histonů může určit stupeň exprese a místní zhutnění chromatinu. Na globálních úrovních je zhutňování chromatinu regulováno také kovalentními modifikacemi histonů v nukleosomech.
Ukázalo se například, že konstitutivní heterochromatin (který není nikdy exprimován a je hustě zabalený) má tendenci ulpívat na jaderné vrstvě a ponechávat jaderné póry volné..
Konstitutivní euchromatin (který je vždy exprimován, například ten, který zahrnuje geny pro udržování buněk a nachází se v oblastech laxního chromatinu), tak činí ve velkých smyčkách, které vystavují transkripci DNA transkripčnímu stroji.
Další oblasti genomové DNA oscilují mezi těmito dvěma stavy v závislosti na době vývoje organismu, podmínkách růstu, identitě buněk atd..
Aby genomy eukaryotických organismů splnily svůj plán pro vývoj, expresi a údržbu buněk, musí jemně regulovat, kdy a jak se musí projevit jejich genetické potenciály..
Vycházejíc z informací uložených v jejich genech, jsou tyto umístěny v jádru v konkrétních oblastech, které určují jejich transkripční stav..
Můžeme tedy říci, že další ze základních rolí nukleosomů, prostřednictvím změn v chromatinu, které pomáhá definovat, je organizace nebo architektura jádra, které je obsahuje..
Tato architektura je zděděna a je fylogeneticky zachována díky existenci těchto modulárních prvků informačního balení.


Zatím žádné komentáře