The buněčné spoje jsou to kontaktní můstky, které existují mezi cytoplazmatickými membránami mezi sousedními buňkami nebo mezi buňkou a matricí. Spojení závisí na typu studované tkáně a zdůrazňuje existující spojení mezi epiteliálními, svalovými a nervovými buňkami.
V buňkách existují molekuly související s adhezí mezi nimi. Ke zvýšení stability vazby v tkáních jsou však zapotřebí další prvky. Toho je dosaženo spojením buněk.
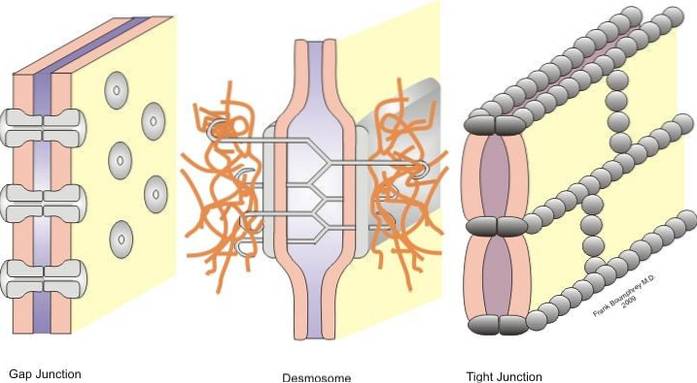
Spojení se dělí na symetrická spojení (těsná spojení, pásové desmosomy a štěrbinová spojení) a asymetrická spojení (hemidesmosomy).
Těsná spojení, desmosomy pásu, bodové desmosomy a hemidesmosomy jsou spoje, které umožňují ukotvení; zatímco rozštěpy se chovají jako spojovací můstky mezi sousedními buňkami, což umožňuje výměnu rozpuštěných látek mezi cytoplazmy.
Pohyb rozpuštěných látek, vody a iontů probíhá prostřednictvím a mezi jednotlivými buněčnými složkami. Existuje tedy transcelulární cesta, která je řízena řadou kanálů a transportérů. Na rozdíl od paracelulární dráhy, kterou regulují kontakty mezi buňkami - tedy buněčné spoje.
V rostlinách najdeme buněčné spoje, které připomínají rozštěpy, nazývané plasmodesmata. I když se liší strukturou, funkce je stejná.
Z lékařského hlediska se určité nedostatky v buněčných spojích promítají do získaných nebo zděděných onemocnění způsobených poškozením epiteliální bariéry.
Rejstřík článků
Živé organismy jsou tvořeny diskrétními a rozmanitými strukturami nazývanými buňky. Ty jsou odděleny plazmatickou membránou, která je udržuje odděleně od extracelulárního prostředí..
I když jsou to složky živých bytostí, nepodobají se cihlám, protože nejsou navzájem izolovány.
Buňky jsou prvky, které jsou ve vzájemné komunikaci a v extracelulárním prostředí. Proto musí existovat způsob, jak buňky mohou vytvářet tkáně a komunikovat, zatímco membrána zůstane neporušená..
Tento problém lze vyřešit díky přítomnosti buněčných spojů, které existují v epitelu. Tyto unie jsou vytvořeny mezi dvěma sousedními buňkami a jsou klasifikovány podle funkce každé z nich v symetrických a asymetrických spojeních.
Hemidesmosomy patří k asymetrickým svazkům a těsné svazky, pásové desmosomy, desmosomy a rozštěpové svazky k symetrickým svazkům. Níže podrobně popíšeme každý z odborů.
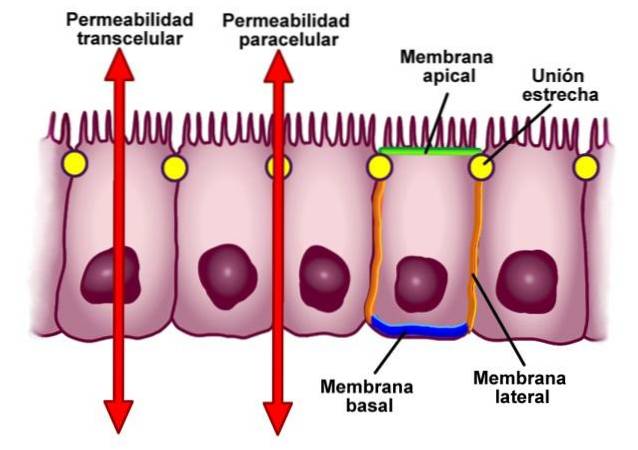
Těsná spojení, v literatuře známá také jako okluzivní spojení, jsou sektory v buněčných membránách sousedních buněk, které jsou úzce propojeny - jak naznačuje název „těsné spojení“.
Za průměrných podmínek jsou buňky odděleny vzdáleností 10-20 nm. V případě těsných spojů je však tato vzdálenost výrazně snížena a membrány obou buněk vedou k dotyku nebo dokonce ke sloučení..
Typický těsný spoj je umístěn mezi bočními stěnami sousedních buněk v minimální vzdálenosti od jejich apikálních povrchů..
V epiteliální tkáni vytvářejí všechny buňky takové spojení, aby zůstaly pohromadě. V této interakci jsou buňky uspořádány do vzoru připomínajícího prsten. Tyto odbory pokrývají celý obvod.
Úzké kontaktní oblasti obklopují celý povrch buňky. Tyto oblasti tvoří anastomované kontaktní proužky transmembránových proteinů známých jako occludin a claudin. Termín anastomóza označuje spojení určitých anatomických prvků.
Tyto dva proteiny patří do skupiny tetraespaninů. Vyznačují se čtyřmi transmembránovými doménami, dvěma vnějšími smyčkami a dvěma relativně krátkými cytoplazmatickými konci..
Bylo prokázáno, že okluzin interaguje se čtyřmi dalšími proteinovými molekulami, které se nazývají zonulový oklucin a zkráceně ZO. Tato poslední skupina zahrnuje proteiny ZO 1, ZO 2, ZO 3 a afadin.
Claudin, na druhé straně, je rodina 16 proteinů, které tvoří řadu lineárních fibril v těsných spojích, což umožňuje tomuto spojení převzít roli „bariéry“ v paracelulární dráze..
Nektiny a molekuly adheze spojení (JAM) se také objevují v těsných spojích. Tyto dvě molekuly se nacházejí v nitrobuněčném prostoru jako homodimery..
Nektiny jsou připojeny k aktinovým vláknům prostřednictvím proteinu afadinu. Ta druhá se jeví jako zásadní, protože při delecích genu, který kóduje afadin u hlodavců, vedou ke smrti embrya.
Tyto typy spojení mezi buňkami plní dvě základní funkce. Prvním z nich je určit polaritu buněk v epitelu, oddělit apikální doménu od bazolaterální domény a zabránit nedovolené difúzi lipidů, proteinů a dalších biomolekul..
Jak jsme uvedli v definici, buňky epitelu jsou seskupeny do kruhu. Tato struktura odděluje apikální povrch buňky od bočního a bazálního, což určuje diferenciaci mezi doménami.
Tato separace je považována za jeden z nejdůležitějších konceptů při studiu fyziologie epitelu..
Za druhé, těsná spojení zabraňují volnému průchodu látek přes vrstvu epiteliálních buněk, což se promítá do bariéry pro paracelulární dráhu..
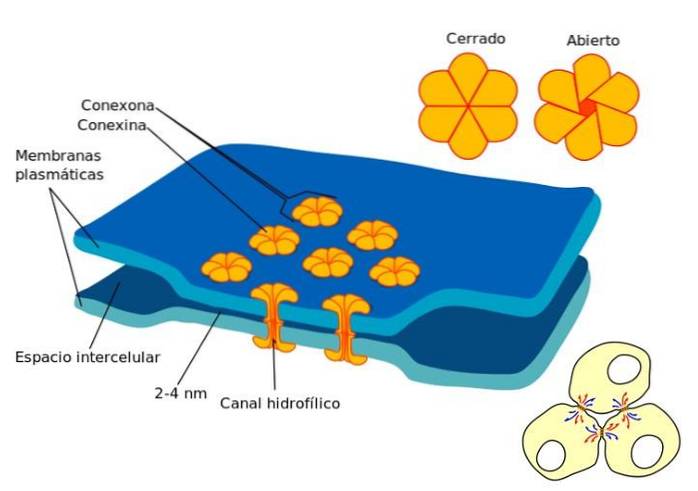
Mezery se nacházejí v oblastech bez omezení cytoplazmatické membrány mezi sousedními buňkami. V rozštěpu se cytoplazmy buněk spojí a vytvoří se fyzické spojení, kde může dojít k průchodu malých molekul..
Tato třída spojů se vyskytuje prakticky ve všech epitelích a v jiných typech tkání, kde slouží k nejrůznějším účelům..
Například v různých tkáních se rozštěpy mohou otevřít nebo zavřít v reakci na extracelulární signály, jako je tomu v případě neurotransmiteru dopaminu. Přítomnost této molekuly snižuje komunikaci mezi třídou neuronů v sítnici v reakci na zvýšenou intenzitu světla..
Rozštěpové spoje jsou tvořeny proteiny zvanými konexiny. Spojení šesti konexinových monomerů se tedy získá „konexon“. Tato struktura je dutý válec, který prochází cytoplazmatickou membránou.
Spoje jsou uspořádány takovým způsobem, že mezi cytoplazmy sousedních buněk je vytvořen kanál. Také spojky mají tendenci se agregovat a tvořit jakési desky..
Díky vytvoření těchto spojů může dojít k pohybu určitých molekul mezi sousedními buňkami. Rozhodující je velikost transportované molekuly, optimální průměr je 1,2, stejně jako ionty vápníku a cyklický adenosinmonofosfát.
Konkrétně se jedná o anorganické ionty a molekuly rozpustné ve vodě, které lze přenést z jedné buněčné cytoplazmy do souvislé cytoplazmy..
Koncentrace vápníku hrají v tomto kanálu zásadní roli. Když se zvyšuje koncentrace vápníku, axiální kanály mají tendenci se uzavírat.
Tímto způsobem se rozštěpy aktivně účastní procesu elektrické a chemické vazby mezi buňkami, k čemuž dochází ve svalových buňkách srdce, které jsou odpovědné za přenos elektrických impulsů..
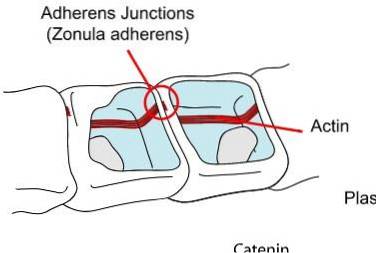
Pod těsnými spoji najdeme kotevní spoje. Obecně se nacházejí poblíž apikálního povrchu epitelu. V této skupině můžeme rozlišit tři hlavní skupiny, zonula adherens nebo pásový desmosom, makula adherens nebo přesný desmosom a desmosom.
V tomto typu spojení jsou sousední buněčné membrány, které jsou spojeny zonulemi a přilnavými makuly, odděleny relativně velkou vzdáleností buněk - ve srovnání s minimálním prostorem, který existuje v případě těsných spojů..
Mezibuněčný prostor zabírají proteiny, které patří do rodiny kadherinů, desmogleinů a desmocholinů navázaných na cytoplazmatické plaky, které představují další proteiny zvané desmoplakin, plakoglobin a plakofilin.
Stejně jako v případě těsných spojů sledujeme v kotevních spojích také vzor uspořádání ve formě prstence nebo pásu. Zonula adherens je spojena s aktinovými mikrofilmenty prostřednictvím interakce dvou proteinů: kadherinů a kateninů..
V některých případech je tato struktura známá jednoduše jako desmosom, jedná se o bodové spojení, které je spojeno s mezivlákny vytvořenými z keratinu. V této souvislosti se tyto keratinové struktury nazývají „tonofilimanetos“. Vlákna se táhnou od bodu k bodu v epitelových buňkách.
Ty poskytují sílu a tuhost buňkám epitelu. Předpokládá se tedy, že jeho hlavní funkce souvisí s posílením a stabilizací sousedních buněk..
Desmosomy lze přirovnat k jakémusi nýtu nebo svaru, protože připomínají oddělené malé tečky a ne spojité pásy.
Tento typ spojení najdeme v interkalačních discích, které se spojují s kardiocyty v srdečním svalu, a v mozkových obloucích, které lemují vnější povrch mozku a míchy..
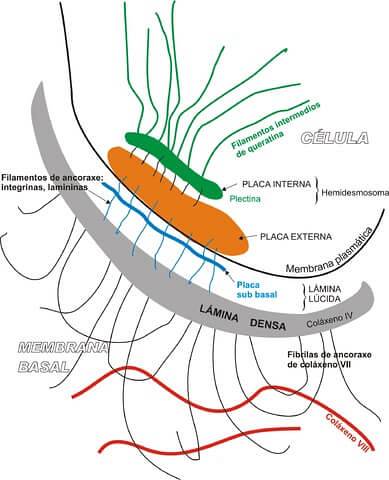
Hemidesmosomy spadají do kategorie asymetrických spojů. Tyto struktury mají funkci ukotvení bazální domény epiteliální buňky se základní bazální laminou.
Termín hemidesmosom se používá proto, že se tato struktura jeví jako doslova „poloviční“ desmosom. Z hlediska jejich biochemického složení jsou však obě unie zcela odlišné..
Je důležité objasnit, že desmosomy jsou odpovědné za adhezi jedné sousední buňky k druhé, zatímco funkcí hemidesmosomu je spojit buňku s bazální laminou.
Na rozdíl od adherenů makuly nebo desmosomu mají hemidesmosomy jinou strukturu, která se skládá z: cytoplazmatické laminy spojené s intermediárními vlákny a destičky vnějších membrán, která je odpovědná za spojení hemidesmosomu s bazální laminou pomocí kotevního vlákna.
Jednou z funkcí hemidesmosomů je zvýšení globální stability epiteliálních tkání díky přítomnosti přechodných cytoskeletálních vláken připojených ke složkám bazální laminy.
V rostlinné říši chybí většina buněčných spojů popsaných výše, s výjimkou funkčního protějšku, který připomíná rozštěpy..
V rostlinách jsou cytoplazmy sousedních buněk spojeny cestami nebo kanály zvanými plasmodesmata..
Tato struktura vytváří kontinuum z jedné rostlinné buňky do druhé. Ačkoli se strukturálně liší od rozštěpových spojů, mají velmi podobné role, které umožňují průchod malých iontů a molekul..
Z lékařského hlediska jsou buněčné spoje relevantní téma. Bylo zjištěno, že mutace v genech kódujících proteiny zapojené do spojení přecházejí do klinických patologií.
Například pokud existuje určitá mutace v genu, který kóduje specifický typ klaudinu (jeden z proteinů, který zprostředkovává interakci v těsných spojích), způsobuje u lidí vzácné onemocnění..
Toto je renální syndrom ztráty hořčíku a příznaky zahrnují nízkou hladinu hořčíku a záchvaty..
Kromě toho bylo zjištěno, že mutace v genu kódujícím protein nektinu 1 je zodpovědná za syndrom rozštěpu rtu. Tento stav je považován za jednu z nejčastějších malformací u novorozenců..
Mutace v genu pro nektin 1 byly také spojeny s dalším stavem zvaným ektodermální dysplázie, který postihuje lidskou kůži, vlasy, nehty a zuby..
Pemphigus foliaceus je puchýřnaté kožní onemocnění určené autoprotilátkami proti desmogleinu 1, klíčovému prvku, který je odpovědný za udržování soudržnosti pokožky.



Zatím žádné komentáře